



Станислав Дробышевский
Палеонтология антрополога. Три эры под одной обложкой
В сланцах Бёрджес сохранились отпечатки Herpetogaster collinsi, сегментированного жирного трёхсантиметрового червяка с растопыренными ветвистыми щупальцами, прикреплённого к почве тоненьким стебельком, внешне среднего между крыложаберными и кишечнодышащими, родственником каковых (и иглокожих тоже) он и являлся; Spartobranchus tenuis – ещё один червяк, сочетающий признаки тех и других, но с перевесом в сторону кишечнодышащих. Другое вероятное полухордовое – Oesia disjuncta, которое первоначально было описано как многощетинковый червь, но одновременно напоминает и щетинкочелюстных.
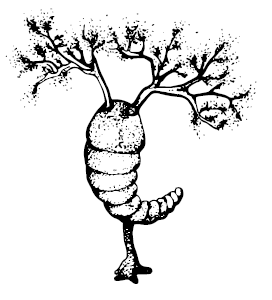
Herpetogaster collinsi
Ещё одна группа нелепых родственников – ветуликолии Vetulicolia; возможно, они тоже были полухордовыми, хотя это не окончательный факт. Древнейшие Vetulicola, Didazoon, Xidazoon и Pomatrum найдены в нижнем кембрии Китая, а Banffia – в Бёрджесе. Передняя часть этих животных представляла собой бочонок или квадратный пакет, покрытый лёгким двустворчатым панцирем, но с жаберными щелями по бокам, а задняя – сегментированную (причём снаружи, что типично для кольчатых червей и членистоногих, но не хордовых) лопасть, служащую, видимо, для гребли, но с кишечником внутри и анальным отверстием на конце. Хорды у них не было, зато на дне глотки имелся эндостиль – желобок с ресничками, типичный для примитивных хордовых. Эти твари были не такими уж мелкими – до 20 см. В среднем кембрии Юты обнаружен другой вариант – Skeemella clavula; передний бочонок похож на ранее найденных, но «хвост» оказался чересчур длинным и откровенно похожим на тело членистоногих.
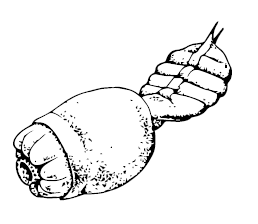
Pomatrum
Промежуточное положение между ветуликолиями, полухордовыми и хордовыми занимают Yunnanozoa (или Yunnanozoonidae, если они не удостаиваются ранга класса, а включаются в какой-то из вышеперечисленных). Самые известные представители – Yunnanozoon lividum и Haikouella jianshanensis из Чэнцзяна и Haikouella lanceolata из Хайкоу. От них остались тысячи отпечатков фантастической сохранности, но это не спасает палеонтологов от сомнений. С одной стороны, юннанозоон и хайкоуелла очень похожи на горбатых ланцетников, у них есть жаберные щели с вроде бы жаберными дугами, вроде бы нотохорд и вроде бы спинной нервный ствол. Однако одновременно вроде бы имеется и брюшной нервный ствол, а в строении кожи, мышц, дыхательной, кровеносной и нервной систем при желании можно найти уйму отличий от идеала хордовых. При более внимательном рассмотрении наличие хорды и жаберных дуг уже выглядит очень сомнительным, а сегментированная кутикула на спине вообще больше напоминает покровы ветуликолий, тогда как нижняя сторона тела вовсе не разделена на миомеры, как бы должно у хордовых. Вокруг рта кругом расположены довольно приличные «зубы», а передний конец увенчан не то круглыми глазами, не то воротничком, аналогичным таковому у кишечнодышащих. В принципе, в такой конфигурации несложно углядеть и черты моллюсков, вероятно, доставшиеся тем и другим от общих ещё вендских предков. Роль юннанозоев в нашей эволюции спорна: одни исследователи считают их первыми позвоночными (только вместо слова «позвоночные» Vertebrata используют термин Cristozoa) и выделяют в группу Procraniata, другие относят их к ветуликолиям или полухордовым.

Banffia
* * *
Весь этот сумасшедший аквариум увенчан настоящими хордовыми Chordata.
Как вы там, потомки?
Самые примитивные представители хордовых – ланцетники Branchiostoma, Epigonichthys и Asymmetron – составляют подтип головохордовых Cephalochordata (или Acrania) с единственным отрядом Amphioxiformes (или Amphioxi). Ланцетники – незамутнённые исходники для всех хордовых, обладают самым лаконичным набором признаков и минимальным количеством специализаций. Хорда, глотка с жаберными щелями, почти замкнутая кровеносная система, нервная трубка, отдельные светочувствительные клетки, печёночный вырост; головы нет (головной конец и голова – не синонимы), головного мозга нет, глаз нет, сердца нет (это надо произносить с надрывом, заламывая руки: «У ланцетника нет сердца!!!»), дыхательной системы нет (жаберные отверстия служат не для дыхания, жабр нет, жаберных дуг нет), скелета нет, черепа нет, конечностей нет. Из оригинальностей – только окружающая жаберные отверстия атриальная полость с отверстием наружу – атриопором, что напоминает мантию моллюсков, а также довольно хитрый ротовой аппарат со щупальцами, парусом (диафрагмой со звездообразным ротовым отверстием между предротовой воронкой и глоткой), эндостилем снизу и наджаберной мерцательной бороздкой сверху. Ланцетник втыкается в землю и фильтрует воду, а если его спугнуть, переплывает подальше и снова втыкается. Довольно долго велись споры: действительно ли ланцетник примитивен, или же он вторично упрощён, а какие-то его предки были вполне даже ничего? Но расшифровка генома расставила всё на свои места: ланцетники правда всегда были такими убогими.
Как ни странно, более продвинуты личиночнохордовые оболочники Urochordata (или Tunicata), включающие асцидий, сальп и аппендикулярий. Асцидии Ascidiacea (или Ascidiae) знамениты разницей в уровнях развития личинок и взрослых особей. Личинки похожи на ланцетника – подвижные, имеют рот, глотку и кишечник (хотя и не питаются), жаберные щели, атриальную полость, хорду (трубка с жидкостью), нервную трубку с мозговым пузырём (что даже круче, чем у ланцетника, причём полость этой трубки отверстием-невропором открыта в ротовую полость) и глазок справа. А вот взрослые асцидии больше похожи на губок или полипов – сидячие, без хорды, зато с двумя сифонами, нервная трубка рассасывается в нервный ганглий; появляется сердце, но капилляров нет, а кровеносная система незамкнутая, выделительная система представлена почками накопления (то есть токсины не выводятся из организма, а скапливаются внутри), кроме полового размножения возможно почкование.
В Куньмине обнаружены отпечатки Shankouclava anningense – древнейших оболочников, очень похожих на асцидий.
Как вы там, потомки?
Сальпы Thaliacea (или Salpae) ещё страннее. Личинки группы бочёночников Doliolida имеют хвост и хорду, но сидят неподвижно в яйце, а у огнетелок Pyrosomida и собственно сальп Salpida их вообще нет. Взрослые же особи подвижны, но с двумя сифонами на концах, а потому двигаются реактивным способом. Вроде бы есть полезные органы – глотка, жаберные отверстия, атриальная полость, желудок, кишка, даже сердце (это куда круче, чем у ланцетника!), но вся прогрессивность хоронится убогим нервным ганглием, незамкнутой кровеносной системой, а особенно – колониальностью и чередованием полового и бесполого размножения. Бесполые особи представляют собой плавающую трубу разного калибра и степени вытянутости (известны огнетелки-пиросомы длиной до 20 м), за которой тянется столон, на общий кишечник которого нанизаны зооиды разного назначения, некоторые из которых отпочковывают самостоятельных половых особей; у огнетелок огромное количество зооидов вогнуто в общий купол-тунику. Когда аквалангисты находят такое чудо, они конечно же тут же снимают его на видео и размещают в интернете с припевами типа «ныряльщики нашли в пучинах инопланетянина!!!» или «открыто неизвестное существо».
И уж совсем запредельно космически выглядят аппендикулярии Appendicularia (или Appendiculariae). По сути своей они являются как бы неотеническими личинками асцидий. Существа эти свободноживущие, но сидят в домике из хитина (вещество, входящее в клеточную стенку грибов и экзоскелет членистоногих), на передней стороне которого есть парные окошечки, затянутые кисеёй из застывшей слизи (сетка быстро засоряется, но это не беда – за минуту-другую аппендикулярия строит новый домик), а сзади – выводное отверстие, реактивной струёй из которого вся конструкция движется. Сама зверюшка внутри домика помахивает длинным хвостом, растущим снизу и спереди, и гонит воду через ловчую сеть, приделанную сзади. В пищеварительной системе, кроме ротовой полости и глотки, предусмотрены желудок и прямая кишка. Как ни странно, у аппендикулярий есть хорда; нервная трубка с мозговым пузырьком личинки превращается в надглоточный ганглий и нервный ствол с ганглиями у взрослой особи; имеется пара жаберных отверстий, но нет атриальной полости; есть сердце, но кровеносная система открытая; органы выделения отсутствуют; почкования нет, но существа гермафродиты, а чтобы всё было совсем уж безумно, личинки разрывают материнскую особь, которая во время «родов» погибает.
Все эти чудеса говорят только об одном: хорда, трубчатая нервная и замкнутая кровеносная системы, подвижность и сообразительность были в кембрии не такими уж очевидно прогрессивными и выигрышными приобретениями. От всего этого можно было запросто отказаться, вернувшись в безмятежность уровня кишечнополостных или даже растворившись в надёжных объятиях собратьев по колонии. Асцидии пошли по пути Обломова, сальпы воплотили идеал «от каждого по способностям, каждому по потребностям». И только самые примитивные и суетные сделали следующий шаг в нашу сторону.
Наконец, мы встречаемся с нашими непосредственными предками – первыми нормальными хордовыми.
В нижнем кембрии Китая обнаружено довольно много кандидатов на звание «проторыб», более продвинутых, чем ланцетник: одно-трёхсантиметровые, сплющенные с боков Cathaymyrus diadexus, C. haikoensis, Zhongxiniscus intermedius и Zhongjianichthys rostratus.

Zhongjianichthys rostratus
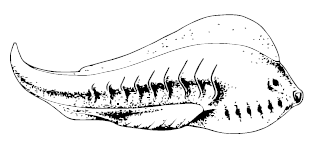
Myllokunmingia fengjiaoa
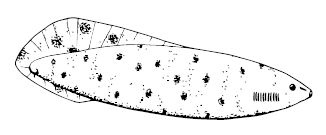
Haikouichthys ercaicunensis
Похожие и очень развитые «проторыбы» – китайские Haikouichthys ercaicunensis и Myllokunmingia fengjiaoa – истинные Великие Предки. Их тело было укреплено нотохордом, окружённым хрящевыми позвоночными элементами, мышцы были подразделены на миомеры, причём в форме шевронов, W-образно, как у ланцетника и рыб, плавниковая складка на спине и хвосте укреплена хрящевыми лучами, на голове имелись жабры с жаберными дугами, большие глаза и даже, вероятно, обонятельные и слуховые капсулы, а в «груди» билось сердце (вряд ли, правда, очень горячее).
В канадском Бёрджесе найдены две подобные «почти рыбы» – Pikaia gracilens и Metaspriggina walcotti. Пикайя более странная: она имела кутикулу, два щупальца спереди и перистые выросты по бокам головного конца, которые похожи на параподии многощетинковых червей, но, скорее всего, были наружными жабрами. Плавниковыми лучами, глазами, сердцем, жаберными дугами и даже межжаберными перегородками она не обладала, миомеры были разделены почти ровными линиями. Даже не факт, что у неё были хорда и нервная трубка, а то, что обычно принимается за таковые, может быть гораздо более мощным «дорзальным органом», возможно, запасающим питательные вещества; тоненькие полосочки под ним, вероятно, всё-таки представляют нервный ствол и нотохорд. Сравнение с более древними, но более продвинутыми китайскими «проторыбами» показывает, что пикайя была специализированной тупиковой версией, альтернативой нашему пути, очевидно, менее удачной.
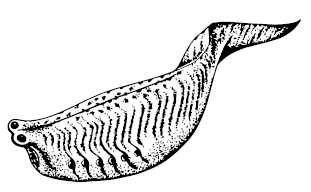
Metaspriggina walcotti
Метасприггина – следующий Великий Предок – гораздо больше похожа на рыбку, у неё есть не только шевропоподобные W-образные миомеры и большие красивые глазки, но и обонятельный орган, и даже зачаток черепа. Её жаберные дуги раздельны, что даже круче, чем у миксин и миног с их жаберной решёткой, а это – первый шаг в сторону обретения челюстей.
* * *
Чего пока не хватало кембрийским хордовым, так это чешуи и зубов. Всякий готов был обидеть беззащитных бедняжек (китайцы и сейчас любят ланцетников в кулинарном смысле, в том числе за то, что их не надо чистить от чешуи, и кость точно не застрянет в горле, так как ни чешуи, ни костей нет), а они не могли ответить как следует. Но дело начало исправляться.
Что появилось раньше – зубы или чешуя, – вопрос философский, так как у древнейших рыбообразных это было примерно одно и то же. Для обменных процессов нужно много фосфора, кальция и фтора, а элементы эти дефицитные; когда находишь их источник, надо не зевать, а срочно лопать и запасать в себе, потому что завтра такого счастья может и не статься. Соли этих элементов твёрдые, при некотором количестве они образуют гранулы, которые, если находятся в коже, становятся чешуёй для защиты, если во рту – зубами для нападения, если внутри тела – костями для приложения мышечной силы. Такие зубчики называются конодонтами Conodonta. Поскольку остальные части тела были мягкими, то понять, как было устроено целое существо, крайне трудно.
Первые конодонтоподобные зубчики – Protohertzina anabarica – известны из самого конца венда – из немакит-далдынского века Алдана в Якутии; P. unguliformis – из томмотского века начала кембрия; их и им подобные относят к протоконодонтам, они очень уж напоминают длинные узкие «клыки» щетинкочелюстных. В то же время в той же Якутии существовали совершенно другие – короткие, с широким основанием и острым кончиком – Fomitchella infundibuliformis («воронковидная»), в Китае Yunnanodus doleres выглядел как шип акации, окружённый пупырышками, а от Западной Европы и Северной Америки (через Якутию и Монголию) до Южного Китая и Австралии жили Lapworthella, похожие на рифлёные бычьи рога. Во второй половине раннего кембрия Rhombocorniculum из Англии и Якутии имели вид длинных бивней, покрытых ажурной ромбической сеточкой, а Mongolodus rostriformis из Монголии – изогнутого когтя. В среднем кембрии появляются вытянутые конусы параконодонты – Amphigeisina, Protohertzina, Gapparodus – и похожие на акульи зубы и подковы истинные эуконодонты – Westergaardodina, Prooneotodus, Proconodontus и прочие. В последующем их становится всё больше и больше, они всё причудливее и причудливее, так что конодонты служат отличными руководящими ископаемыми всего палеозоя. Парадокс в том, что описаны тысячи вариантов, а кем были владельцы этих «зубов» – неведомо. Предполагается, что большая часть конодонтов – действительно зубы хордовых, но явно не все. Уже упоминалось, что конодонтоносители виваксия и одонтогрифус оказались многощетинковым червём и моллюском, Rhombocorniculum – шипами галлюцигений, а когтеподобные Mongolodus – и правда когтями онихофор.
В начале позднего кембрия появляются настоящие рыбообразные, очевидно, пока бесчелюстные. На что они были похожи, науке пока неизвестно, но в Северной Америке найдены чешуйки, описанные как Anatolepis heintzi. Принадлежность анатолеписа именно к рыбам доказывается гистологически. Замечательно, что этот род протянул до среднего ордовика, в это время он известен со Шпицбергена. Судя по однообразию находок, можно предположить, что анатолепис был истинным Великим Предком всех более поздних рыбообразных. Впрочем, другие рыбьи чешуйки, не получившие никакого названия, обнаружены в позднекембрийских отложениях долины Георгины в западном Квинсленде в Австралии. Они имеют своё гистологическое своеобразие, так что можно догадываться, что разделение рыб на группы началось ещё в среднем кембрии. Впрочем, крайняя немногочисленность видов говорит сама за себя.
* * *
Как положено любому приличному периоду, кембрий завершился мощным вымиранием. Во второй половине кембрия фиксируется замедление видообразования, а в конце исчезло 42 % видов морских организмов, в основном брахиопод, трилобитов и конодонтов. Мы точно не знаем причин этого коллапса. С одной стороны, резкое похолодание – уже достаточный повод вымерзнуть, появление полярных ледников и понижение уровня океана – тоже веская причина помереть, с другой, не меньший вклад могли внести новые хищники, например, головоногие наутилоидеи. Так или иначе, исчезновение множества прежних властителей океана открыло дорогу новым формам. Наступал ордовик.
* * *
В кембрии мы обрели генеральный план строения своего тела: голова с парой глаз и обонятельными рецепторами спереди-снизу, на спинной стороне – нервная трубка с головным мозгом из пяти мозговых пузырей на переднем конце, опорно-двигательный аппарат из сегментированных мышц, поддерживаемый остовом в центре, на брюшной стороне – пищеварительная трубка, передняя часть которой заодно занимается дыханием; довершает картину замкнутая кровеносная система с сердцем. В общем-то, за вычетом ручек и ножек, кембрийские предки – это уже готовые мы.
Альтернативы
«Кембрийский взрыв» предоставил широчайшие возможности жизни. Огромное количество альтернативных линий наперебой занимали все возможные экологические ниши. Донные и планктонные, фильтраторы и хищники, бронированные и мягкие, умные и водоросли – все они стартовали примерно одновременно и имели условно равные шансы. Самые разнообразные черви могли увеличиться, нарастить нервную систему, вооружиться зубами, ускориться плавниками и стать царями природы. Будущее могло быть за ксенузиями и онихофорами, супер тихоходками и аномалокаридами, трилобитами и наутилоидеями. Каждая из этих и многих других групп имела свои плюсы и преимущества. Бурное изобилие челюстей и ног, коготков и зубов, панцирей и стилей жизни, варящееся в горячем кембрийском океане – тот бульон, из которого неизбежно должны были выйти более совершенные существа. Тем печальнее, что многие из подававших большие надежды быстро достигли идеала и остановились в развитии, застыли, сохранив свой кембрийский облик доныне. Как бы выглядели разумные потомки приапулид или ветуликолий? К чему бы они стремились, какие цели перед собой ставили?
Ордовик
485,4–443,8 миллиона лет назад: Мир наоборот
МЕЖДУНАРОДНАЯ И РОССИЙСКАЯ ШКАЛА:
485,4 млн л.н. – ранний ордовик: тремадокский век – 477,7 – флоский век – 470 – средний ордовик: дапинский век – 467,3 – дарривильский век – 458,4 – поздний ордовик: сандбийский век – 453 – катийский век – 445,2 – хирнантский век – 443,8

Ордовик – не очень длинный, но богатый на живых существ период; с другой стороны, какой-то одной, самой впечатляющей и харизматичной, присущей именно ордовику группы как-то не нашлось. На роль таковых могли бы претендовать трилобиты и наутилоидеи, но они существовали и до, и после ордовика. Впрочем, и ордовику есть чем похвастаться.
* * *
География ордовика прямо обратна современной. На Южном полюсе зависла Гондвана, слепленная из Африки, Южной Америки, Австралии и Антарктиды, так что почти строго на полюсе лежали Центральная Африка и Амазония. Вдоль экватора выстроились Северная Америка и кусочки Евразии, так что самыми экваториальными областями оказались Сибирь и полярная Канада с Гренландией. Северная Европа плыла по Южному полушарию на широте нынешней Австралии. На просторах Центральной Азии – Якутии, Тувы, Алтая и монгольской пустыни Гоби – плескалось тёплое тропическое море. Странные времена, странные карты.
Как бы там ни было, материки в очередной раз начали сближаться, а климат, соответственно – резко холодать, так что от кембрийского максимума к концу периода он установился до почти современных значений, лишь на четыре градуса теплее нынешнего. Впрочем, вплоть до середины периода климат был тёплым и влажным, способствовавшим химическому выветриванию и, соответственно, почвообразованию. Впрочем, о наземных экосистемах ордовика мы почти ничего не знаем. Другое дело – моря.
В ордовике произошёл один из самых мощных рывков разнообразия живых организмов за всю историю планеты, число семейств выросло более чем вдвое; при этом большая часть кембрийских форм тоже продолжала успешно существовать. Сопоставимые преобразования случались только в кембрии (но тогда возникли вообще все, с кембрием соревноваться бессмысленно), в юре-меле и кайнозое, но для них у нас намного больше данных.
Из растений в ордовике бурный расцвет переживали красные водоросли Rhodophyta и зелёные ульвофициевые, или сифоновые водоросли Ulvophyceae (или Siphonophyceae), появившиеся ещё как минимум в кембрии. Красные водоросли ордовика, представленные древней группой Solenoporaceae, и сифоновые водоросли часто имеют известковый покров, благодаря чему неплохо сохраняются в ископаемом виде.
Как вы там, потомки?
Сифоновые водоросли уникальны тем, что их тело представляет собой недослившуюся-недоразделившуюся одну гигантскую многоядерную клетку. Особенно показательны Caulerpa и Acetabularia, похожие чаще на поганки с тонюсенькой ножкой, но иногда – на перья или банановые листья. Некоторые виды достигают огромных размеров: многоядерные клетки Acetabularia mediterranea имеют длину 18 см, а Caulerpa prolifera – более 3 м!
Работа над ошибками
Типовой вид Solenopora spongioides, давший название всей группе Solenoporaceae, до крайности похож на губку и изначально – ещё в 1877 году – так и был определён. Ровно через полвека, в 1927 году, на его основе была выделена группа примитивнейших красных водорослей, образцом которых соленопора считалась ещё много десятилетий. Однако в 2004-м колесо палеонтологической сансары сделало свой полный оборот, и соленопора опять была признана губкой. Впрочем, это не отменяет водорослевую природу всех других соленопоровых ордовика и силура.
В начале ордовика появляются кораллы табуляты Tabulata, в начале среднего ордовика они дают великое разнообразие. В отличие от современных кораллов, табуляты так и не стали настоящими рифостроителями. Они жили на сравнительно небольшой глубине, часто в компании с брахиоподами и кольчатыми червями, но редко – с моллюсками, которые предпочитали более глубокие участки. С начала среднего ордовика к табулятам добавились четырёхлучевые кораллы ругозы Rugosa. Эти часто были одиночными, в виде расширяющегося наверх конуса на тонкой ножке, причём очень крупными и тяжёлыми, так что, вырастая, они падали на бок и продолжали жить лёжа; колониальные же, как и родственники табуляты, не строили значимых рифов.
В раннем ордовике расцветают мшанки Bryozoa (они же Ectoprocta) – странная группа животных, альтернативная линия эволюции беспозвоночных.
Как вы там, потомки?
Внешне мшанки выглядят крайне просто – мелкие полипчики со щупальцами, обычно собранные в мохообразные колонии, покрывающие камни на дне. Но у мшанок есть вторичная полость тела и сквозная пищеварительная система, а также нервный ганглий, хотя бы и один, так что они – потомки каких-то гораздо более совершённых животных, чем кишечнополостные. Личинки мшанок крайне разнообразны: билатеральные, некоторые с двойной раковинкой, обычно покрытые ресничками; широко распространено и почкование, и формирование особых покоящихся статобластов для переживания плохих времён. Самое удивительное свойство мшанок – колониальность, причём особи, формирующие колонию, имеют разное строение. Колонии обычно неподвижные, но бывают и ползающие, внешне похожие на голожаберных моллюсков. Кроме стандартных питающих автозооидов с лофофором из щупалец, есть индивиды оеции – выводковые камеры, специальные кенозооиды для прикрепления к субстрату, гонозооиды для полового размножения, защитники – клювастые авикулярии с открывающейся вниз «челюстью» и вибракулярии, трясущие здоровенным шипом. Общий известковый скелет колонии может быть защищён всяческими пупырышками и шипами. И вот это последнее свойство дало мшанкам возможность строить с начала ордовика целые мшанковые рифы, на которых губки и кораллы играли подчинённую роль. Мшанки могли захватить мир! Но, как часто бывает, более примитивные кишечнополостные всё же взяли верх; впрочем, это было уже после ордовика.
Ближайшие родственники мшанок – брахиоподы тоже достигли в ордовике несказанного изобилия, но… оставаясь при этом внешне совершенно невыразительными и однообразными. Восхищаться бесконечными видами, родами и семействами брахиопод может только полный фанатик.
Особое величие обрели головоногие моллюски Nautiloidea. Их главным достижением было совершенствование прямой или слабо изогнутой раковины, внутри перегороженной на камеры, заполненные газом. Такая конструкция позволяла наутилоидеям подниматься и опускаться по вертикали, а потом разгоняться реактивным способом, выбрасывая воду из мантийной полости через сифон. Спереди из раковины высовывалась голова. В Северной Америке найдены следы в виде расположенных полукругом подковок – какой-то подобный моллюск шарил по илистому дну извивающимися короткими щупальцами. Среди наутилоидей предельных размеров достигли Endocerida (или Endoceratoidea): раковины среднеордовикских Endoceras или Cameroceras (эти два рода различимы с трудом) вырастали от 3,5 до 6 м длиной (раньше самые крупные оценивались даже в 9,5 м, но потом цифры были пересмотрены). Это почти рекорд длины для животных не только ордовика, но и вообще палеозоя!
Обычно эндоцериды представляются как крупнейшие хищники своего времени. Но такой образ имеет много проблем. Во-первых, эндоцериды были маломанёвренными, они здорово двигались по прямой, но не могли быстро поворачивать. Во-вторых, несмотря на многочисленность находок, никто никогда не находил их челюстей, тогда как потенциальная добыча – трилобиты – были неплохо защищены панцирями. В-третьих, обкусанных трилобитов удивительно мало, несравнимо с обилием раковин эндоцерид. В-четвёртых, эндоцериды какие-то больно щуплые – при двухметровой длине диаметр раковины достигал всего десятка сантиметров. Наводит на мысли о какой-то специфической специализации и резкое вымирание группы в конце периода. Кроме прочего, распределение находок свидетельствует о том, что взрослые крупные особи держались над большими глубинами. По итогу, эндоцериды получаются не кровожадными хищниками, а поедателями всякой мелкой планктонной мути, болтавшейся в толще воды. Вероятно, они плавали, растопырив щупальца с натянутой на них перепонкой, как это делают многие современные глубоководные осьминоги типа Vampiroteutis infernalis (оцените харизматичное название!), и отцеживали из воды мелких рачков. Случившийся в конце ордовика планктонный кризис подорвал кормовую базу несчастных головоногих – и они исчезли.
Эндоцериды – самая характерная для ордовика группа: они возникли и вымерли в пределах этого периода, в раннем силуре доживали свой век лишь единичные их представители. Впрочем, другие прямораковинные родственники, например Actinocerida (они же Actinoceratoidea), прожили намного дольше – до начала карбона, Bactritida (или Bactritoidea) – до начала позднего триаса, а Orthocerida (иногда вместе с несколькими родственными отрядами выделяемые в свой подкласс Orthoceratoidea) – и вовсе до самого конца триаса.
Как вы там, потомки?
Наутилус Nautilus – далёкий потомок ордовикских наутилоидей, обрётший кругло-спиральную раковину с чудесной полосатой рыже-белой раскраской, живёт в тропической части запада Тихого океана и в наши дни. Это настоящее «живое ископаемое»; наутилус сохранил массу примитивных черт древнейших головоногих: очень примитивные глаза в виде колбы, открывающейся отверстием наружу, без хрусталика, роговицы и стекловидного тела, четыре жабры, четыре почки, четыре сердца, воронка сифона сделана из двух сложенных, но не сросшихся лопастей, щупальца чрезвычайно многочисленны и лишены присосок, есть широкая радула, а чернильной железы нет. Главное приобретение наутилуса по сравнению с палеозойскими предками – мощные роговые челюсти, которыми можно щёлкать креветок.
Не исключено, что древние прямораковинные наутилоидеи тоже были красивыми, по крайней мере раковины Proterovaginoceras belemnitiforme сверху были тёмно-бурыми, а снизу светлыми.
В нижнеордовикских слоях Марокко сохранились отпечатки аномалокарид, достигавших в длину, вероятно, больше метра. По крайней мере, они превосходят более древних канадских родственников, хотя, возможно, уступают некоторым китайским.
Среди настоящих членистоногих продолжали развиваться некоторые экзотические группы. Самая известная позднеордовикская Marrellomorpha — Furca bohemica – чудное существо, похожее не то на мчащуюся морскую звезду с шестью полощущимися на ветру мохнатыми лучами, не то на наскальный рисунок вислорогого снежного человека. Как ни странно, членистых ног, на сохранившихся отпечатках, вообще не видно. Вероятно, они были полностью спрятаны под шестилучевым панцирем или вовсе атрофировались.
Работа над ошибками
Первые отпечатки фурки были приняты за ротовой аппарат трилобита, потом переопределены как личинки иглокожих и лишь намного позже опознаны как полноценные самостоятельные существа.
Ордовикские трилобиты переживали эволюционный успех. Про них написаны отдельные толстые книги, здесь мы скажем лишь о самых выдающихся представителях. Isotelus rex из позднего ордовика Канады – крупнейший трилобит, достигавший длины 72 см; учитывая его крайне коренастую комплекцию и полное отсутствие выступающих шипов, он был самым массивным созданием ордовика! Родственный вид Isotelus brachycephalus был вдвое скромнее, но тоже не карликом – 33 см. Другой позднеордовикский гигант – испанский Uralichas hispanicus tardus – вырастал до 70 см, хотя некоторую часть его длины составлял хвостовой шип. Марокканский Declivolithus alfredi имел почти дисковидную форму за счёт разросшегося в виде широкой пластины головного щита, окружающего всё тело и покрытого множеством ямок. Шестисантиметровый Illaenus умел сворачиваться в идеальный шар благодаря выпуклым началу и концу и укороченной середине. Cyclopyge – мелкий трилобитик, тоже способный превращаться в сферическую консерву, с огромными глазами, занявшими почти всю круглую голову (впрочем, вопреки названию, так и не слившимися в один), бодро плававший в толще воды.
Особенно многочисленны и разнообразны трилобиты ордовика Ленинградской области: Ampyx nasutus, Lonchodomas rostratus, L. volborthi и Glopampyx linnarssoni имели на головном щите три тончайших и длиннющих выроста – один торчащий вперёд из «носа» и два по бокам, закинутые назад намного дальше конца тела; у Hoplolichas plautini колючки короткие, зато их много, как у ехидны; у Hoplolichoides furcifer над спиной тянулись два толстых рога, как у антилопы, у Paraceraurus macrophtalmus были два изогнутых назад дугами рога – отростка головного щита – и два саблевидных рога-хвоста; Boedaspis ensifer впечатляет ещё больше – он во все стороны ощетинился длинными изогнутыми «рогами» – выростами всех сегментов. На последнего похож североамериканский Ceratocephala laciniata. Вся эта красота появилась явно неспроста: защита не возникает на пустом месте. Очевидно, трилобиты очень нравились каким-то хищникам, имевшим прочные челюсти для разгрызания панцирей.
Были и другие способы избавиться от напастей. Asaphus kowalewskii, судя по длинным глазным стебелькам, глубоко закапывались в ил. Как и кембрийские оленусы, нижнеордовикские трилобиты Cloacaspis и позднеордовикские Triarthrus заняли экстремальную, но относительно безопасную нишу передвижных плантаций для симбиотических серобактерий. Правда, периодически из-за падения уровня кислорода они сами вымирали большими массами, но за отсутствие конкурентов и хищников тоже надо платить. Несмотря на примитивность, некоторым трилобитам, видимо, не были чужды зачатки социального поведения, о чём свидетельствуют цепочки забавных марроканских Ampyx priscus с коротеньким тельцем и длиннющими «носами» и шипами. Порой в такие колонны встраивались совершенно непохожие виды, например, Asaphellus jujuanus. Зачем они ходили строем, прикасаясь друг к другу – загадка, есть много версий, из которых основная – миграция.




