



Е. И. Николаева
Психофизиология. Психологическая физиология с основами физиологической психологии. Учебник
Электроэнцефалограмма и методы ее регистрации
В 1929 г. австрийский психиатр Ганс Бергер обнаружил, что с помощью игольчатых платиновых электродов, помещенных на различные точки поверхности головы человека, можно зарегистрировать электрическую активность мозга. Эти записи он и назвал электроэнцефалограммой (ЭЭГ). Хотя ЭЭГ снимается с поверхности головы, Г. Бергер сумел доказать, что часть электрической активности обусловлена деятельностью мозга, а не покрывающих его поверхность тканей (Berger, 1929).
Открытие Г. Бергера было встречено весьма холодно, и этот метод получил признание лишь после того, как Е. Д. Эдриан и Б. Х. Мэттьюз смогли непосредственно продемонстрировать его на заседании Английского физиологического общества в 1935 г. Сам Е. Д. Эдриан был в качестве испытуемого и, закрывая глаза, показал появление альфа-ритма на ЭЭГ (Хэссет, 1981).
Интерпретация электроэнцефалографических данных требует учета целого ряда параметров – формы волн, их амплитуды, расположения электродов, с которых получена запись, и т. д. Наиболее важными из них являются амплитуда волн и частота. Амплитуда измеряется как расстояние от базовой линии до пика волны. Однако существуют значительные трудности в определении базовой линии, поэтому на практике обычно используют измерение от пика до пика (рис. 2.3). Под частотой понимается число полных циклов, совершаемых волной за 1 сек. Этот показатель измеряется в герцах (Гц). Величина, обратная частоте, называется периодом волны. Известно, что амплитуда волн ЭЭГ находится в обратно пропорциональной зависимости от их частоты (Cooper e. a., 1980).
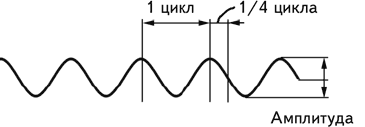
Рис. 2.3. Амплитуда волны и ее цикл.
Ритмичность электроэнцефалографического сигнала позволяет количественно описать записи ЭЭГ. Первым из описанных ритмов электрической активности мозга был альфа-ритм (Berger, 1929). Он представляет собой ритмические колебания электрической активности 8–12 раз в сек (8–12 Гц) с амплитудой около 50 мкВ. Эта активность преобладает у здорового человека в состоянии спокойного бодрствования с закрытыми глазами и наблюдается преимущественно в затылочных областях (рис. 2.4). Альфа-ритм отсутствует у слепорожденных людей (Новикова, 1978). Ритмами той же частоты, но с иной формой волны и в специфических отведениях, являются мю-ритм, регистрируемый в роландовой борозде, каппа-ритм, отмечаемый при наложении электродов в височном отведении (рис. 2.5).
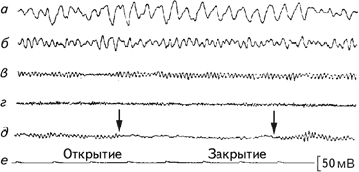
Рис. 2.4. Классификация волн ЭЭГ по частоте. Образцы дельта- (а), тета- (б), альфа- (в), и бета- (г) активности; (д) – блокада альфа-ритма при открытии глаз; (е) – отметка времени (Cooper, 1980).
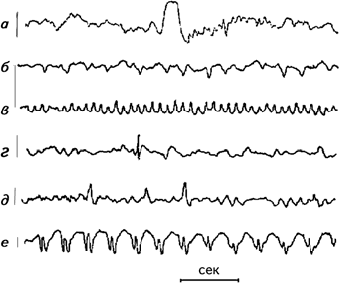
Рис. 2.5. Образцы специфических волн: а – К-комплекс; б – ламбда-волна; в – мю-ритм; г – спайк; д – острая волна; е – комплекс спайк-медленная волна. Слева дается калибровочная линия, соответствующая 100 мВ, справа – расположение электродов на голове. Внизу отметка времени 1 сек (Cooper, 1980).
Г. Бергер описал и другой тип волн – бета-ритм, обнаруживаемый у человека в состоянии активного бодрствования. Это колебания с частотой более 13 Гц и небольшой (около 25 мкВ) амплитудой.
Дальнейшие исследования выявили более медленные колебания, которые были названы тета- и дельта-волнами, а также более быстрые – гамма-волны. Тета-волны имеют частоту от 3,5 до 7,5 Гц и амплитуду от 5 до 100 мкВ, дельта-волны – 1–3,5 Гц и 20–200 мкВ соответственно. Чем медленнее волны, тем больше их амплитуда. Гамма-волны – колебания с частотой более 30 Гц и амплитудой около 2 мкВ (рис. 2.4).
Каждый из ритмов имеет свою преимущественную локализацию: альфа-ритм регистрируется в затылочном и теменном отведениях; тета-волны – в лобных и височных, бета-ритм – в прецентральных и фронтальных, гамма-ритм – в прецентральных, фронтальных, височных, теменных; дельта-ритм не имеет определенной локализации.
При визуальном анализе ЭЭГ обычно определялись два показателя – длительность альфа-ритма и блокада альфа-ритма, которая фиксируется при предъявлении испытуемому того или иного раздражителя (рис. 2.4). Экспериментаторы также пытались обнаружить на ЭЭГ испытуемых особые волны, отличающиеся от фоновой регистрации. Даваемое им название связано либо с их формой, либо с тем местом, где они выявляются. К ним относятся: К-комплекс, ламбда-волны, мю-ритм, спайк, острая волна.
К-комплекс – это сочетание медленной волны с острой волной, вслед за которыми часто идут волны частотой около 14 Гц. К-комплекс возникает во время сна или спонтанно у бодрствующего человека. Максимальная амплитуда отмечается в вертексе и обычно не превышает 200 мВ (Roth e. a., 1956) (рис. 2.5). Ламбда-волны – монофазные положительные острые волны, возникающие в окципитальной области, связанные с движением глаз. Их амплитуда меньше 50 мВ, частота – 12–14 Гц (Green, 1957). Мю-ритм – группа аркообразных или гребневидных волн частотой 7–11 Гц, амплитудой меньше 50 мВ, регистрирующихся в центральных областях головы. Они блокируются двигательной активностью или тактильной стимуляцией (Gastaut e. a., 1952). Спайк – волна, четко отличающаяся от фоновой активности, с выраженным пиком длительностью от 20 до 70 мс. Первичный компонент обычно является негативным (Maulsby, 1971). Острая волна – ясно отличающаяся от фоновой активности волна с подчеркнутым пиком длительностью 70–200 мс. Спайк-медленная волна – последовательность поверхностно негативных медленных волн (частотой 2,5–3,5 Гц), каждая из которых ассоциируется со спайком. Амплитуда этого комплекса может достигать 1000 мВ (Chatrian e. a., 1968) (рис. 2.5).
Визуальный анализ позволяет оценить лишь очевидные изменения биоэлектрической активности мозга (рис. 2.6). В настоящее время разработан метод для регистрации медленных и сверхмедленных потенциалов, имеющих длительность периода от нескольких секунд и более (Илюхина, 1977). Такие потенциалы регистрируются с использованием усилителей постоянного тока (рис. 2.7).
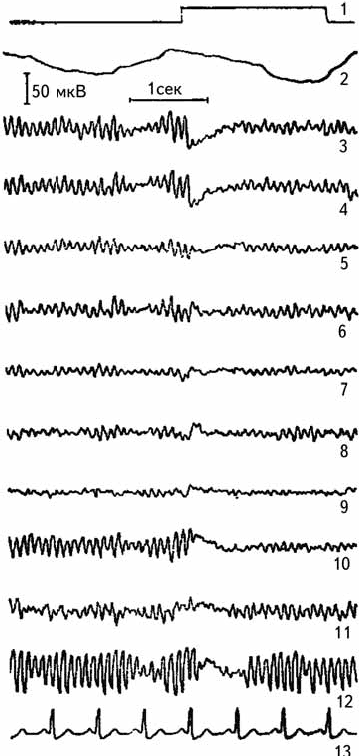
Рис. 2.6. Реакция депрессии альфа-ритма при непрерывном освещении глаза. 1 – отметка раздражителя, 2 – пневмограмма, 3–12 – униполярная регистрация ЭЭГ, 13 – ЭКГ (Лебедева, 1977).
Для более глубокого анализа используются компьютерные методы обработки сигнала, позволяющие извлекать существенно больше информации. Традиционный способ интерпретации электрических сигналов, отраженных в нерегулярных временных рядах, основан на разложении этих сигналов с помощью Фурье-преобразования. При этом в сигнале выделяются частотные компоненты. Затем рассчитывается мощность ЭЭГ в каждой частотной полосе для всех отведений, и с помощью специальных цветовых шкал результаты представляются в виде наглядных картин, где цвет отражает интенсивность ритмов каждого диапазона в различных частях мозга. Этот метод получил название картирования мозга (рис. 2.8). Возможно также использование не цвета, а изолиний при изображении одних и тех же мощностей показателя.
Однако спектральный анализ ЭЭГ в этой парадигме имеет ограниченные возможности в формировании картины электрической активности мозга. В настоящее время появилась возможность применять для анализа ЭЭГ нелинейную теорию систем (Lutzenberger e. a., 1992).
При нелинейном динамическом анализе исследуемые сигналы ЭЭГ рассматриваются как стационарные, то есть предполагается, что значения средних, дисперсии и автоковариационная функция остаются неизменными в течение определенного промежутка времени. Реальные сигналы ЭЭГ сохраняют стационарность в течение длительных интервалов времени с небольшой вероятностью, поэтому эта проблема решается несколькими способами. Прежде всего, для анализа используются относительно короткие временные промежутки (эпохи). Считается, что ошибка в абсолютной оценке параметров за столь короткий срок будет систематической, что позволит корректное сравнение контрольного и экспериментального условий (Pritchard, Duke, 1995). Второй способ решения проблемы заключается в использовании методов, нечувствительных к нестационарности (Skinner, 1994).
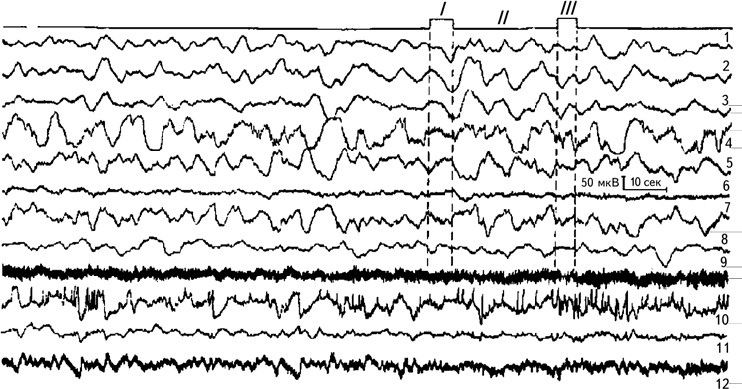
Рис. 2.7. Динамика медленных электрических процессов в подкорковых образованиях головного мозга человека при тесте на оперативную память.
I – предъявление теста; II – этап сохранения; III – воспроизведение теста. 1, 2, 3 – вентролатеральное ядро зрительного бугра; 4 – скорлупа; 5, 6, 7 – бледный шар; 8 – красное ядро; 9 – внутренняя капсула; 10 – передневентральное ядро зрительного бугра; 11 – ретикулярное ядро зрительного бугра; 12 – срединный центр зрительного бугра. Регистрация на энцефалографе с постоянной времени 2,5 сек (Илюхина, 1977).
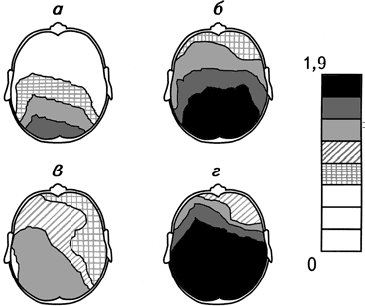
Рис. 2.8. Суммарные топографические карты амплитуды спектра мощности α-ритма ЭЭГ при прослушивании классической (а, б) и рок-музыки (в, г).
а, в – здоровые испытуемые, б, г – больные депрессией. Из каждой карты вычтена соответствующая ей фоновая карта. Справа – шкала амплитуд спектра мощности, мкВ (Михайлова, 1992).
Чтобы отделить сигнала от шума, требуется высокое качество регистрируемого сигнала, либо из кусков качественных записей конструируются наборы суррогатных (искусственных) временных рядов, что позволяет сравнивать результаты нелинейного динамического анализа для оригинальных данных и фильтрованного шума. Несмотря на отсутствие однозначных доказательств действительной хаотичности реального сигнала ЭЭГ, механизм его генерации указывает гипотетически на происхождение от нелинейной динамической системы, управляемой квазипериодическими законами (Афтанас, 2000).
Для характеристики нейрональных динамических систем используются две принципиально различные категории нелинейных динамических мер. К первой категории относится размерностная комплексность (синонимы: корреляционная размерность, фрактальная размерность). Она является показателем общей комплексности (сложности) корковой динамики и отражает ее статические свойства (Pritchard, Duke, 1995). Для изучения динамических характеристик системы, ее изменений во времени, используют энтропию Колмогорова (К2), наибольшую экспоненту Ляпунова (L1) и нелинейное предсказание. Энтропия Колмогорова оценивает степень хаотичности системы, то есть среднюю скорость, с которой информация о системе теряется. Если этот показатель стремится к нулю, то есть нет изменений информации о системе, система становится полностью предсказуемой. Наоборот, для хаотичной системы метрика энтропии имеет определенное конечное положительное значение. И чем больше эта величина, тем более хаотичной является система. Спектр экспонент Ляпунова дает оценку схождения или расхождения близлежащих траекторий в фазовом пространстве, отражая чувствительность системы к исходным значениям. Наличие положительной главной экспоненты является дополнительным свидетельством хаотического поведения изучаемой системы (Афтанас, 2000). Нелинейное предсказание или прогноз характеризует вероятность, с которой может быть предсказана динамика временного ряда. Она основывается на расчете коэффициентов корреляции между наблюдаемым и предсказанными временными рядами. Поэтому она позволяет отслеживать потерю информации во времени.
Впервые оценка фрактальной размерности ЭЭГ произведена в 1985 г. (Babloyantz e.a., 1985). Сейчас получены доказательства относительно более низкой размерности альфа-ритма по сравнению с суммарной ЭЭГ (Pritchard, Duke, 1995).
Динамическая мера фрактальной размерности отражает минимальное количество активированных нейронов и нейрональных цепей, лежащих в основе исследуемого сегмента ЭЭГ. Синхронизация их активности отражает состояние покоя корковых сетей и соответствует высокой степени кооперации между нейронными сетями, что обнаруживается в низкой фрактальной размерности. Десинхронизация, связанная с процессами активной переработки информации в коре головного мозга, характеризует усиление конкурентного взаимодействия между нейронными сетями и сопровождается повышением фрактальной размерности ЭЭГ. Снижение конкуренции между клеточными ансамблями (и, следовательно, снижение уровней фрактальной размерности ЭЭГ) может происходить не только вследствие синхронизации корковых процессов, но и в результате более эффективного использования специализированных для данного типа переработки информации областей коры. В этом случае наблюдается выключение активности нейронных сетей, не вовлеченных в выполнение данного задания, в результате чего более низкие уровни фрактальной размерности ЭЭГ могут отражать интенсификацию процессов переработки информации вовлеченными областями коры.
Другим способом анализа процессов, происходящих в ЦНС, является метод пространственной синхронизации биопотенциалов мозга или когерентности. Этот метод был предложен и разработан М. Н. Ливановым (1972; 1989). Когерентный анализ позволяет определить сходство активности различных областей коры в одном и том же частотном диапазоне и, предположительно, оценить количество корко-корковых соединений и их интенсивность между двумя областями коры (Ливанов, 1989).
Оценка пространственной синхронизации биопотенциалов мозга, основанная на быстром преобразовании Фурье, получила дальнейшее развитие в работах А. М. Иваницкого, предложившего новый метод картирования внутрикорковых взаимодействий. Он основывается на выявлении в ЭЭГ с помощью спектрального анализа пиков активности, совпадающих в разных отведениях по частоте (Иваницкий, 1990, 1996).
В настоящее время разработана новая нелинейная мера – совместная фрактальная размерность, позволяющая количественно оценить степень динамического взаимодействия двух одновременно регистрируемых аттракторов из различных областей коры головного мозга. (Stam e.a., 1996). Каждый из этих аттракторов представляет собой одновременно включенные в процесс самоорганизованные в корковом пространстве нейронные сети, осциллирующие на различных частотах (Stam e.a., 1996). Оценка совместной размерности ЭЭГ не просто отражает фрактальную размерность в области индивидуального отведения, а характеризует степень динамического кооперативного взаимодействия между пространственно разделенными аттракторами. Это принципиально отличает механизмы совместной размерности от процессов, которые отражаются в когерентности, поскольку когерентность оценивает степень взаимодействия дистантно расположенных нейронных сетей, осциллирующих на одной частоте (Ливанов, 1989; Иваницкий, 1996).
Динамика здоровой физиологической системы должна продуцировать высоко нерегулярные и комплексные типы вариативности, в то время как заболевание и старение связаны с потерей комплексности и большей регулярностью (Ehlers, 1995).
Запись ЭЭГ осуществляется с помощью разнообразных электродов, форма и исполнение которых зависит от специфики исследования. Они должны обладать малым переходным сопротивлением, минимальной поляризацией и антикоррозийными свойствами. Среди наиболее распространенных материалов, используемых для электродов, – серебро, посеребренная медь, графит (Гречин, 1977). Подпаянные к электродам проводники, необходимые для подключения датчиков на вход приборов, должны обладать высококачественной изоляцией. Для улучшения фиксации ЭЭГ используют разнообразные прокладки и пасты, снижающие переходное сопротивление, а кожа испытуемого предварительно обезжиривается спиртом или смесью спирта с эфиром (50 х 50).
Для того чтобы результаты различных исследователей можно было сопоставлять, Международная федерация обществ ЭЭГ рекомендовала стандартный метод расположения электродов на поверхности скальпа – систему «10–20» (Jasper, 1958). В соответствии с требованиями этой системы производят три основных измерения: расстояние по сагиттальной линии между носовой впадиной и затылочным бугром, длину от одного наружного слухового прохода через макушку головы до другого во фронтальной плоскости, длину окружности головы. Каждая из этих величин принимается за 100 %. В продольном направлении лобный и затылочный электроды устанавливаются в точках, отстоящих от переносицы и затылочного бугра на расстоянии 10 % общего расстояния. Остальные 3 электрода располагаются на расстояниях 20 % от этих двух и друг от друга.
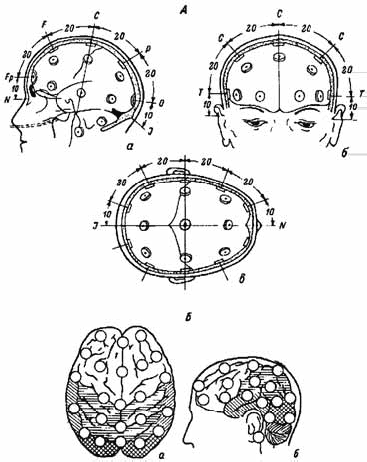
Рис. 2.9. Международная схема расстановки электродов – схема «10–20».
А: а – вид сбоку, б – вид спереди, в – вид сверху (сечение по височной линии). Б – проекция электродов на доли мозга: а – вид сверху, б – вид сбоку. Косая штриховка – височная область, перекрестная – затылочная, горизонтальная – теменная область; не заштрихована лобная область и область центральной извилины. N – переносица. J – затылочный бугор (Jasper, 1958).
Всего устанавливается 5 рядов электродов: один по сагиттальной линии и по два параллельных ряда справа и слева на равных расстояниях, составляющих по 20 % от общей длины во фронтальной плоскости. Височные ряды электродов отстоят от наружных слуховых проходов на 10 % общего поперечного расстояния. В общей сложности на поверхности головы располагается 21 электрод (рис. 2.9).
Записи ЭЭГ являются информативными при диагностике эпилепсии или мозговых опухолей, исследовании стадий сна и бодрствования (рис. 2.10).
Являясь достаточно эффективным методом в оценке активности мозга в целом, электроэнцефалография не позволяет судить о возбуждении отдельных нейронов или нейронных ассоциаций.
Регистрация импульсной активности нервных клеток
Импульсная активность отдельных нейронов может оцениваться лишь у животных и в отдельных случаях во время оперативного вмешательства на мозге человека. Впервые активность коркового нейрона головного мозга человека была зарегистрирована А. Уардом и Л. Томасом (Ward, Thomas, 1955) в процессе операции у больного по поводу эпилепсии.
Сейчас для регистрации нейронной импульсной активности головного мозга человека используются микроэлектроды с диаметром кончиков 0,5–10 мкм. Они могут быть выполнены из нержавеющей стали, вольфрама, платиноиридиевых сплавов, золота. Электроды вводятся в мозг с помощью специальных микроманипуляторов, позволяющих точно подводить электрод к нужному месту.
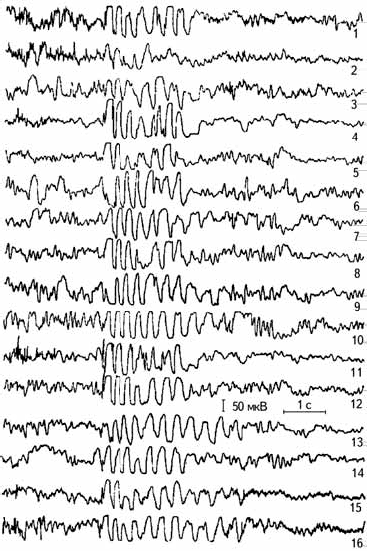
Рис. 2.10. Провокация пароксизмальной активности при гипервентиляции (Лебедева, 1977). 1–16 – каналы отведений
С 1964 г. производят регистрацию импульсной активности нейронов подкорковых структур мозга при стереотаксических операциях. Стереотаксический аппарат позволяет вводить электрод с большой точностью в необходимую структуру, расположенную в глубине мозга (Трохачев, Матвеев, 1977).
В некоторых случаях больным вживляют долгосрочные электроды, например, для выявления эпилептогенных очагов. В отличие от одномоментной регистрации применение электродов, оставляемых на длительный срок в ткани мозга, имеет свои особенности. Возможность стабильной регистрации появляется только на 15–20-й день после операции, что связано с морфологическими изменениями мозгового субстрата, возникающими из-за введения инородного тела – электрода, и сохраняется в течение 3–5 месяцев.
В этом случае речь идет о внеклеточном (экстрацеллюлярном) отведении потенциалов. Если электрическая активность отдельного нейрона имеет определенный ритм, закономерно изменяющийся при его различных функциональных состояниях, то электрическая активность группы нейронов обладает сложной структурой. На нейрограмме в этом случае отражается суммарная активность многих нейронов, возбуждающихся в разное время, различающихся по амплитуде, частоте и фазе (рис. 2.11). Полученные данные обрабатываются автоматически по специальным программам (Трохачев, Матвеев, 1977).
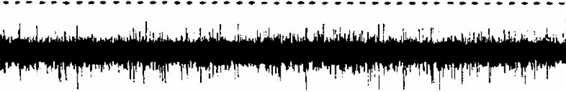
Рис. 2.11. Электрограмма нейронного пула. Вверху штрихами произведена отметка времени м/сек (Трохачев, Матвеев, 1977).
Регистрация вызванных потенциалов мозга человека и потенциалов, связанных с событиями
Предъявление одиночного стимула мало что меняет в картине ЭЭГ, поскольку эти изменения маскируются общей фоновой активностью. Для того чтобы сделать изменения в ответ на определенный стимул видимыми, используют компьютер, усредняющий записи волновой активности при повторных предъявлениях того же стимула. Случайная по отношению к предъявлению стимула электрическая активность нивелируется, тогда как специфическая активность, связанная со стимулом, будет усиливаться. Такая активность называется вызванным потенциалом (ВП). Этот биоэлектрический феномен был описан еще в XIX веке Ричардом Кейтоном (Caton, 1877, см.: Петушков, 1977). Технику когерентного накопления волновой активности при повторных предъявлениях стимула разработал в 1951 г. Дж. Доусон. В зависимости от поставленных в исследовании задач усреднение ЭЭГ активности производилось либо относительно стимула, либо от начала движения, следовавшего за стимулом. Полученные в различных экспериментальных ситуациях потенциалы стали называть общим термином – потенциалы, связанные с событиями.
Вызванный потенциал (ВП) состоит из последовательности отрицательных и положительных отклонений от основной линии и длится около 500 мсек после окончания действия стимула. У ВП можно оценить амплитуду и латентный период возникновения (рис. 2.12).
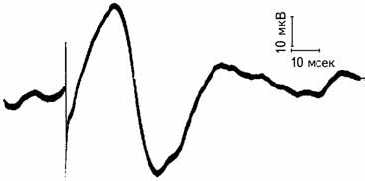
Рис. 2.12. Одиночный вызванный потенциал. Регистрация с поверхности головы в области проекции сенсомоторной зоны мозга при электрической стимуляции латерального отдела зрительного бугра (Jasper, 1958).
На величину ВП может влиять модальность стимула. Так, слуховые ВП отличаются от зрительных, зрительные – от тактильных и т. д. Вызванные потенциалы, возникающие в ответ на световые, звуковые, тактильные или электрические раздражения в проекционной зоне соответствующего анализатора и имеющие короткий латентный период, называются первичными; все остальные, обладающие другим пространственным распределением и большим латентным периодом, – вторичными, или поздними ответами (Петушков, 1977).
Один и тот же ВП может быть обусловлен многими психологическим процессами, а одни и те же психические процессы могут быть связаны с разными ВП (Event… 1991).
Для регистрации ВП используются те же электроды, что и для записи ЭЭГ, и требуется соблюдение определенных условий проведения исследований. К ним относится унификация методических приемов всей серии экспериментов, проведение их на одном и том же испытуемом, в одно и то же время, в одном и том же состоянии, с использованием одних и тех же параметров стимуляции. Обработка данных включает в себя методы математического, статистического анализа. В простейшем случае определяют латентный период, амплитуду и длительность отдельных фаз (Петушков, 1977).