



Алексей Макарушин
Парадоксы эволюции. Как наличие ресурсов и отсутствие внешних угроз приводит к самоуничтожению вида и что мы можем с этим сделать
Библиографический список
1. Селье Г. (1982). Стресс без дистресса. – М: Прогресс.
2. ФГБУН «ФИЦ Питания, биотехнологии и безопасности пищи» (2020). Оценка влияния регулярного употребления воды с высоким содержанием бикарбоната натрия на анаэробную производительность. Отчет о научно-исследовательской работе.
3. Александер Д., Райхлен Д. (2020). Зачем мозгу физические упражнения? ВМН, № 3, с. 5–11.
4. Понцер Г. (2017). Парадокс физической активности. ВМН, № 4, с. 27–33.
5. Понцер Г. (2019). Эволюция и тренировки. ВМН, № 3, с. 5–13.
6. Шубин Н. (2008). Внутренняя рыба. История человеческого тела с древнейших времен до наших дней. – М.: Династия.
7. Radak Z., Chung H. Y., Koltai E., Taylo, A. W., Goto S. (2007). Exercise, oxidative stress and hormesis, Ageing Res. Rev. 170.
8. Malm C., Sjodin T. L., Sjoberg B., Lenkei R., Renstrom P., Lundberg I. E., Ekblom B. (2004). Leukocytes, cytokines, growth factors and hormones in human skelet al. muscle and blood after uphill or downhill running. J. Physiol. 556, 983–1000.
9. Peake J. M., Nosaka K., Muthalib M., Suzuki K. (2006). Systemic inflammatory responses to maximal versus submaximal lengthening contractions of the elbow flexors. Exerc. Immunol. Rev. 12, 72–85.
10. Goto S., Radak Z. (2005). Proteins and exercise. In: Mooren, F. C., Volker, K. (Eds.), Molecular and Cellular Exercise Physiology, Human Kinetics. Champaign, USA, 55–71.
11. Nieman D. C., Wentz L. M. (2019). The compelling link between physical activity and the body’s defense system. J Sport Health Sci; 8: 201–17.
12. Radak Z., Ishihara K., Tekus E., Varga C., Posa A., Balogh L., Boldogh I., Koltai E. (2017). Exercise, oxidants, and antioxidants change the shape of the bell-shaped hormesis curve. Redox Biology 12, 285–290.
13. Windsor M. T., Bailey T. G., Perissiou M., Meital L., Golledge J., Russell F. D., Askew C. D. (2018). Cytokine Responses to Acute Exercise in Healthy Older Adults: The Effect of Cardiorespiratory Fitness. Front. Physiol. 9: 203.
14. Fischer C. P. (2006). Interleukin-6 in acute exercise and training: what is the biological relevance? Exercise Immunology Review, 6–33.
15. Leal L. G., Lopes M. A., Batista M. L. (2018). Physical Exercise-Induced Myokines and Muscle-Adipose Tissue Crosstalk: A Review of Current Knowledge and the Implications for Health and Metabolic Diseases. Front. Physiol. 9: 1307.
16. Northoff H., Berg A. (1991). Immunologic mediators as parameters of the reaction to strenuous exercise. Int J Sports Med 12, Suppl, 1: S, 9–15.
17. Larrabee R. C. (1902). Leukocytosis after violent exercise. J Med Res (NS); 7: 76–82.
18. Kishimoto T. (1989). The biology of interleukin-6. Blood 74: 1–10.
19. Kerksick С. М., Wilborn C. D., Roberts M. D., Smith-Ryan A., Kleiner S. M., Jäger R., Collins R., Cooke M., Davis J. N., Galvan E., Greenwood M., Lowery L. M., Wildman R., Antonio J., Kreider R. B. (2018). ISSN exercise & sports nutrition review update: research & recommendations. Journal of the International Society of Sports Nutrition 15: 38.
20. Chycki J., Kostrzeva M., Maszczyk A., Zajac A. (2021). Chronic ingestion of rich-bicarbonate water improves anaerobic performance in hypohydrated elite combat sport athletes. Int J of Env Res and Public Health, 18.
21. Nicholls D. G., Bernson V. S., Heaton G. M. (1978). The identification of the component in the inner membrane of brown adipose tissue mitochondria responsible for regulating energy dissipation Experientia Suppl.,Vol. 32., 89–93.
22. Rupprecht A., Sittner D., Smorodchenko A., Hilse K. E., Goyn J., Moldzio R., Seiler E., Anja U., Brauer A. E. M., Pohl E. E., (2014). Uncoupling Protein 2 and 4 Expression Pattern during Stem Cell Differentiation Provides New Insight into Their Putative Function. PLoS ONE 9(2): e88474.
23. Chaudhuri L., Srivastava R. K., Kos F., Shrikant P. A. (2016). Uncoupling protein 2 regulates metabolic reprogramming and fate of antigen-stimulated CD8+ T cells, Cancer Immunol Immunother. 65 (7): 869–874.
24. Echtay K. S., Winkler E., Bienengraeber M., Klingenberg M. (2000). Site-directed mutagenesis identifies residues in uncoupling protein (UCP1) involved in three different functions. Biochemistry; 39: 3311–3317.
25. Dietrich M. O., Andrews Z. B., Horvath T. L. (2008). Exercise-Induced Synaptogenesis in the Hippocampus Is Dependent on UCP2-Regulated Mitochondrial Adaptation. Journal of Neuroscience 15 October, 28 (42): 10766–10771.
26. Raschke S., Elsen M., Gassenhuber H., Sommerfeld M., Schwahn U., Brockmann B., Jung R., Wisløff U., Tjønna A. E., Raastad T., Hallén J., Norheim F., Drevon C. A., Romacho T., Eckardt K., Eckel J. (2013). Evidence against a Beneficial Effect of Irisin in Humans//PLoS ONE. – September 11 (Vol. 8, № 9), e73680.
27. Raichlen D. A., Klimentidis, Y. C., Bharadwaj, P. K., Alexander G. E. (2019). Differential Associations of engagement in physical activity and estimated cardiorespiratory fitness with brain volume in middle-aged to older adults in Brain Imaging and Behavior.
28. Raichlen D. A., Alexander G. E. (2017). Adaptive capacity: an evolutionary neuroscience model linking exercise, cognition, and brain health. Trends in Neurosciences, Vol. 40, № 7, 408–421.
29. Clark A., Mach N. (2016). Exercise-induced stress behavior, gut microbiota-brain axis and diet: a systematic review for athletes. Journal of the International Society of Sports Nutrition, 13–43.
30. Zeppa S. D., Agostini D., Gervasi M., Annibalini G., Amatori S., Ferrini F., Sisti D., Piccoli G., Barbieri E., Sestili P., Stocchi V. (2020). Mutual Interactions among Exercise, Sport Supplements and Microbiota. Nutrients 2020, 12, 17.
31. Oettlé G. J. (1991). Effect of moderate exercise on bowel habit. Gut, 32, 941–944.
32. Hagio M., Matsumoto M., Yajima T., Hara H., Ishizuka S. (1985). Voluntary wheel running exercise and dietary lactose concomitantly reduce proportion of secondary bile acids in rat feces. J. Appl. Physiol., 109, 663–668.
33. Cerda B., Perez M., Perez-Santiago J. D., Tornero-Aguilera J. F., Gonzalez-Soltero R., Larrosa M. (2016). GutMicrobiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health, Front. Physiol., 7, 51.
34. Francaux M. (2009). Toll-like receptor signalling induced by endurance exercise. Appl. Physiol. Nutr. Metab., 34, 454–458.
35. Frosali S., Pagliari D., Gambassi G., Landolfi R., Pandolfi F., Cianci R. (2015). How the Intricate Interactionamong Toll-Like Receptors, Microbiota, and Intestinal Immunity Can Influence Gastrointestinal Pathology. J. Immunol. Res., 2015, 489821.
36. Viloria M., Lara-Padilla E., Campos-Rodríguez R., Jarillo-Luna A., Reyna-Garfias H., López-Sánchez P., Rivera-Aguilar V., Salas-Casas A., Berral de la Rosa F.J., García-LatorreE. (2011). Effect of moderate exercise on IgA levels and lymphocyte count in mouse intestine. Immunol. Investig., 40, 640–656.
37. Turnbaugh P. J., Ley R. E., Mahowald M. A., Magrini V., Mardis E. R., Gordon J. I. (2006). An obesity-associated gut microbiome with increased capacity for energy harvest. Nature, 444, 1027–1031.
38. Mailing L. J., Allen J. M., Buford T. W., Fields C. J., Woods J. A. (2019). Exercise and the Gut Microbiome: A Review of the Evidence, Potential Mechanisms, and Implications for Human Health. Exerc. Sport Sci. Rev. 2019, 47, 75–85.
39. Dokladny K., Zuhl M. N., Moseley P. L. (2016). Molecular Adaptations to Exercise, Heat Acclimation, and Thermotolerance. J. Appl. Physiol. 15; 120 (6): 692–701.
40. Andrews P. W., Thomson J. A. Jr. (2009). The bright side of being blue: depression as an adaptation for analyzing complex problems. Psychol. Rev.; 116 (3): 620–54.
41. Stix G. (2021). Evolution Could Explain Why Psychotherapy May Work for Depression. Scientific American, February, 14.
42. Clark A., Mach N. (2017) The Crosstalk between the Gut Microbiota and Mitochondria during Exercise. Front. Physiol. 8: 319.
43. Lutz H. L., Jackson E. W., Webala P. W., Babyesiza W. S., Kerbis Peterhans J. C., Demos T. C., Patterson B. D., Gilbert J. A., (2019). Ecology and host identity outweigh evolutionary history in shaping the bat microbiome. mSystems, 4: e 00511–19.
Глава V. Цель – все и ничто
Только хорошо поняв второе начало термодинамики, можно ответить на вопрос, почему вообще возможна жизнь.
Карло Черчиньяни (1998), итальянский математик, физик-теоретик, поэт и эссеист
Энтропия к месту и не очень
Разумеется, в эволюционном движении живого, как разновидности движения материи и энергии, нет никакого предопределенного смысла, а тем более замысла – ни скрытого, ни тем более явного, несмотря на то что они «ясно видны» многим «посвященным». Если где-то в данной книге и используются выражения вроде «эволюция сделала вот это, чтобы решить вот то», на самом деле это всего лишь «обытовленное» объяснение динамики генетических ансамблей, к которым «принципы статистической физики применимы столь же, сколь и к ансамблям молекул» (Кунин Е. В., 2012).
Все формы взаимодействия в биологических и протобиологических средах (сложных органических молекул) можно в своих крайних пределах свести к двум формам: отношениям протагонистов или дружественности (сотрудничество, синтрофия, симбиоз, кооперация и т. д.) и отношениям антагонистов или враждебности (прямое соперничество, конкуренция, с одной стороны, и хищничество, паразитизм, зачастую вырастающий из сотрудничества, с другой). Граница между ними тонкая, если не зыбкая; нейтральных же взаимодействий, пожалуй, нет совсем. Соответственно, ответ на вопрос, на каких этапах развития материи и в какой последовательности эти явления возникают, и каким образом они вытекают из оснований статистической физики, может стать и предвестником ответа на вопрос о сущности болезней, воспринимаемых часто как потери совершенства изначально почти безупречных организмов или как прямые следствия их некоего изначального несовершенства. Представляется, однако, что именно несовершенства, как отклонения от совершенного низкоэнтропийного «идеала», реальные или кажущиеся, и делают живой (и не только живой) мир таким, каков он есть: развивающимся от простого к все более сложному, постоянно генерирующим что-то новое, неизбежно забывающим и неизбывно повторяющим (почти) все старое, в котором все в отдельности неизбежно скоро заканчивается и когда-то закончится и все целиком.
В третьей главе на отдельных примерах были кратко рассмотрены квантовые и энтропийные основания базовых энергетических предпосылок жизни и сопутствующая им диссипативность, то есть «набор свойств открытой, неравновесной, рассеивающей энергию системы, движимой градиентом энергии, что увеличивает энтропию окружающей среды». Несомненно, что физические, в первую очередь термодинамические основания, второе начало термодинамики являются наиболее всеобъемлющим оправданием существования всего живого с его общим движением к нарастанию энтропии, но с неизбежным возникновением локальных и преходящих ее снижений (флуктуаций), то есть упорядочиваний хаоса. Рассмотрение энтропии как чистой «меры беспорядка» может запутать: очевидно, что энтропия «покоящейся» системы – ровного песка в пустыне, сбалансированной группы клеток, стагнирующего человеческого общества – гораздо выше, соответственно, энтропии осыпающегося бархана, смеси клеток хозяев и клеток новых паразитов, общества в состоянии революции и гражданской войны, ввиду того, что совокупность или возможность «выбора» доступных состояний для каждой единицы или вариантов совокупностей микросостояний в «спокойной» системе гораздо больше возможностей выбора в состоянии «революции».
Так, лидер партии «Яблоко» Григорий Явлинский в своей статье «Политическая энтропия. Цифровые технологии и глобализация беспорядка» (2020) предполагает, что на смену эпохи глобализации, предположительно, более упорядоченной, приходит «век беспорядка», где «существует множество труднопредсказуемых событий и развилок, и куда, как говорится, кривая выведет, не знает никто». И этот переход с ростом беспорядка предполагается рассматривать как рост «политической энтропии». Однако более правильным будет скорее обратное – падение энтропии: несмотря на возникновение потоков событий с непредсказуемой динамикой, для отдельных людей – совокупности элементов системы – количество вариантов выбора («свободы») на самом деле уменьшается, о чем совершенно справедливо указывает и сам Г. Явлинский в заключение своей статьи, связывая это в первую очередь с добровольным отказом людей от свободы выбора, нежели направленной деятельностью авторитарных лидеров. Отдельный большой вопрос, разумеется, насколько этот выбор на самом деле доброволен, но несомненно, что само уменьшение свободы выбора для подавляющего большинства элементов системы – это несомненно снижение ее энтропии. Тем более актуальное в свете взрывного развития информационных технологий и роста все более детализированной информации о системе. Информации, к сожалению, неравномерно распределяемой, но в итоге дающей уменьшение неполноты информации о системе (см. ниже), что также показывает скорее падение «политической энтропии», нежели ее возрастание. И тем более, что «добровольный отказ от свободы» и рост информационной насыщенности общественного пространства могут оказаться двумя сторонами одной медали.
Термодинамика и информация
Со времен установления принципа Людвига Больцмана (ок. 1875 г.) стало возможным рассматривать классическую термодинамическую и информационную энтропию как практически и теоретически очень близкие, если не равнозначные. В формулировке Макса Планка (ок. 1900 г.) этот принцип как связь средней энтропии системы с вероятностью данного состояния определяется логарифмической зависимостью:
S = kln(Ω),
где S – общая энтропия состояния системы, константа k = = 1,38*10-23 Дж/К, названная Планком постоянной Больцмана, а Ω – статистический вес состояния, являющийся числом возможных равновероятных микросостояний (способов), с помощью которых можно составить данное макроскопическое состояние. Более ранняя формулировка Джозайя Гиббса (1878 г.) для микросостояний, имеющих разную вероятность, устанавливает зависимость в виде суммы произведения вероятности состояний на натуральные логарифмы этих же вероятностей, взятой с противоположным знаком:
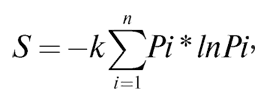
где Pi – вероятность пребывания системы в состоянии i. Минус в начале формулы необходим для приведения обеих частей формулы к общему знаку из-за постоянной отрицательности логарифмов вероятности (так как вероятности всегда меньше 1) и неизменной положительности энтропии. Сам Больцман, комментируя свое физико-статистическое определение энтропии, отмечал, что энтропия характеризует недостающую информацию о системе. В этой интерпретации энтропия может считаться мерой неполноты информации о системе.
В ходе разработки углубленной теории информации информационной энтропией стала считаться мера неопределенности, независимо выведенная как сумма произведений вероятностей событий на логарифмы этих же вероятностей (формула Хартли-Шеннона, основателей теории информации), взятая с противоположным знаком:
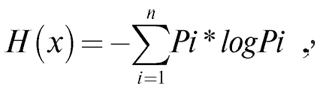
где H(x) – средняя энтропия сообщения, иногда обозначаемая также как мера неопределенности какого-либо события, или даже как количество информации, Pi – вероятность отдельного события i. Основание логарифма в принципе может быть любым больше 1: в двоичных информационных системах – 2, математические статистики, использующие формулу в различных научных дисциплинах, в том числе биологических и клинических, часто используют натуральный логарифм.
Отрицательная энтропия, которую описывал Эрвин Шредингер, это, разумеется, не энтропия с отрицательным знаком – это, очень условно говоря, мера удаленности от максимальной энтропии в рассматриваемой системе, хотя само понятие негативной энтропии (негэнтропии) на самом деле гораздо более сложное, глубокое и весьма запутанное, даже в сравнении с часто неоднозначно понимаемой «обычной», «позитивной» энтропией в своих самых разнообразных интерпретациях. Некоторые исследователи, вслед за Норбертом Винером, увидевшим за очевидным сходством формул Больцмана-Гиббса и Хартли-Шеннона более глубокую связь термодинамической и информационной энтропий, ассоциируют негэнтропию с информацией, что представляется красивым, но до сих пор далеко не всеми признаваемым суждением.
Одной из точек смыкания термодинамической и информационной энтропии являлся, например, известный парадокс «демона Максвелла», гипотетического устройства, стоящего на границе двух объемов и способного различать быстрые и медленные молекулы и, соответственно, пропускать или не пропускать молекулы в одну сторону, что спонтанно ведет к увеличению упорядоченности системы и падению энтропии, что формально противоречит второму началу термодинамики. Решение парадокса было предложено французским и американским физиком Леоном Бриллюэном, математически показавшем, что даже демон ничего не хочет делать бесплатно: чтобы увидеть скорость молекулы, ему нужно заплатить энергией, которая не может быть менее энергии одного теплового кванта (0,025 эВ при комнатной температуре), и эта энергия соответствует минимальной цене одного бита информации. То есть демон совершает работу по обработке информации, и эта работа охлаждает одно тело и нагревает другое, и так же, как за работу любого домашнего холодильника, за эту работу надо платить.
Теория семантической информации
Демон Максвелла получает (увеличивает) информацию о системе, равную уменьшению энтропии (или увеличению негэнтропии), и, казалось бы, отсюда информация = негэнтропия. Однако Бриллюэн различал свободную информацию (без четкого определения) и связанную, возникающую, когда возможные случаи могут быть представлены как микроинформация физической системы, и замечал, что «только связанная информация будет представляться связанной с энтропией».
В российской литературе и части зарубежной со связанной информацией Бриллюэна наиболее соотносится термин «микроинформация», отличающийся от «макроинформации» (практически всегда соответствующей общему термину «информация» в большинстве резонов его применения) тем, что не обладает свойством фиксируемости (запоминаемости). В работах Дэвида Уолперта (см. ниже) с микроинформацией может быть сопоставлена «синтаксическая информация», а с макроинформацией – «семантическая (смысловая) информация». Помимо запоминаемости вторым критерием информации в наиболее оптимальном для практического применения определении информации по Генри Кастлеру является случайность: «информация есть случайный и запомненный выбор одного варианта из нескольких возможных и равноправных» (Кастлер Г., 1967). Важнейший момент в данной теории информации следует из ее определения: возникновение (генерация) информации – это случайный, но запомненный выбор. Второй важный момент – это наличие ценности информации, мерилом которой служит степень ее соответствия цели системы. Она может определяться как функция, включающая отношение вероятности достижения цели (или уменьшения энергетических, временнЫх или материальных затрат, если цель безусловно достижима) после получения информации, к этой вероятности или снижению затрат без обладания этой информацией (Чернавский Д. С., 2004). До получения информации вероятность достижения цели определяется тем, какой предварительной запомненной информацией (тезаурусом) уже располагает принимающий ее элемент. Имея минимумом информации, хотя бы минимальной запомненной информацией «о самом себе», элемент располагает минимальным тезаурусом.
В новой математической теории семантической информации Дэвида Уолперта и Артемия Колчинского (Artemy Kolchinsky and David H. Wolpert, 2018) эта мысль развивается с учетом ключевого влияния окружающей среды: семантическая («макро-») информация определяется как синтаксическая («микро-») информация, которую физическая система располагает о своей окружающей среде, и которая ситуативно (casually) необходима системе, чтобы поддерживать свое существование во времени (что созвучно понятию ценности информации).
Семантическая информация поддерживает длительное существование системы за счет минимизации энтропии системы. Дэвид Уолперт разделяет два вида семантической информации: сохраненную семантическую информацию, которая основывается на обмене общей информацией между системой и окружением в некий исходный момент времени, и наблюдаемую информацию, которая основывается на переносе энтропии между системой и окружением за время от исходного до момента наблюдения. В отличие от синтаксической микроинформации семантическая информация, располагая критерием ценности, может быть ценной (положительной), малоценной или вовсе ошибочной, то есть отрицательной, наносящей урон системе, сокращающей время ее существования. Семантическая информация, в противоположность синтаксической, по Уолперту, фундаментально ассиметрична в ходе любого обмена ею. Другое ключевое понятие, вводимое Уолпертом, это понятие «автономного агента» – физическая система способна быть автономным агентом в той степени, в какой она располагает объемом семантической информации. Данной теорией предполагается, что способность быть живым – это способность быть чрезвычайно развитым автономным информационным агентом.