



Алексей Макарушин
Парадоксы эволюции. Как наличие ресурсов и отсутствие внешних угроз приводит к самоуничтожению вида и что мы можем с этим сделать
Если к горячим источникам добавить цикличность
В последние годы исследовательская группа Тары Джокич, Дэвида Димера и Мартина ван Кранендонка (Tara Jokic, David Deamer and Martin van Kranendonk, 2017) активно разрабатывает альтернативную теорию наиболее вероятного места и механизма происхождения жизни. Они располагают это сакральное место в окрестностях систем древних наземных вулканических водоемов, похожих на современные гейзерные поля Йеллоустоуна или Камчатки, но обладавших цикличностью высыхания-гелеобразования-увлажнения. Подобная цикличность позволяет образовываться многослойным протоорганическим формированиям, предшественникам органических полупроницаемых мембран – первоначально, в сухую фазу, плоским, и изредка во влажную фазу, со сферическими отпочкованиями. Однако энергетический баланс протоклеточных структур в этом случае с трудом покрывается простыми неорганическими источниками энергии, например полифосфатами. Принципиально эта теория не отвергает роль щелочных гидротермальных источников: на примере Боржоми мы видим, что такие источники вполне могут быть наземными; более того, в историческом плане они могут быть наземно-приморскими: та же местность Боржоми еще в позднем миоцене, возможно, представляла собой прибрежную зону деградирующего океана Тетис с разворачивающейся вулканической деятельностью. Вероятность подобных сочетаний в эоархее, предполагаемой эпохе возникновения настоящих жизненных форм, в общем-то тоже не исключена. В подобных случаях цикличность может включать два типа влажных фаз (или серий фаз): щелочную гидротермальную и кислую морскую (приливную?). В любом случае крайне маловероятно, что даже в «чистой» теории щелочных гидротермальных источников линия разграничения щелочного и кислого потоков будет стабильна; более вероятно, что в части микроячеистой породы с большей проницаемостью попеременно меняется рН среды, а в части микроячеек с меньшей проницаемостью рН более стабильно, но меняется их окружение. Возможная «сухая» фаза способствует дополнительной концентрации органических молекул на первичных минеральных мембранах, дополнительно к феноменам термофорезаи компартментализации (то есть концентрирования в пограничных сегментах за счет тепловых градиентов и тепловых конвекционных потоков во множестве полупроницаемых разграничений). В принципе, как показано исследовательской группой под руководством Джулиана Тэннера и Андерсона Шума из Университета Гонконга на примере термодинамики «высыхающих капель», компартментализации в протобиологических структурах может способствовать даже процесс частичного испарения в двухфазовых водных системах (Guo W. et al., 2021).
В последующем неорганическая основа может почти полностью «вымываться» или механически, или кислотным воздействием, подобно вытравливанию кислотой элементной платы, оставляя функциональные каталитические фрагменты – те же железосерные кластеры. С другой стороны, «чистая» теория щелочных гидротермальных источников «выталкивает» наиболее вероятных прародителей биологической изменчивости и наследственности – это неустойчивые в щелочной среде полимеры рибонуклеиновой кислоты (РНК) – на кислую сторону первичных полупроницаемых мембран, где этой теорией не предполагается (хотя и не исключается) ячеистая неорганическая мембранная структура (компартментализация). В комбинированном циклическом варианте с двумя – кислой и щелочной – влажными фазами фрагменты РНК могут быть заключены в ячейку с кислой средой внутри, находящуюся в щелочном окружении. Более того, подобным образом могут организовываться вторичные и третичные матрешкообразные ячеистые формирования, где мелкие кислые пузырьки в окружении более крупных щелочных конгломератов, оказывающиеся внутри крупных кислых кластеров, частично окруженных щелочной средой и так далее (рис. 2–6).
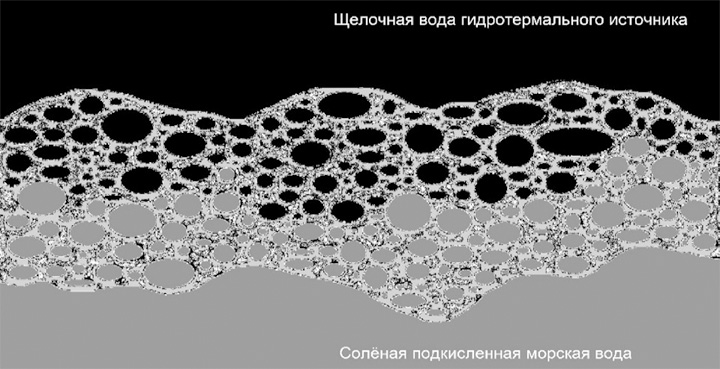
Рис. 2. Микропористые ячейки, проводящие щелочную воду гидротермального источника, частично заполнены также подкисленной соленой морской водой
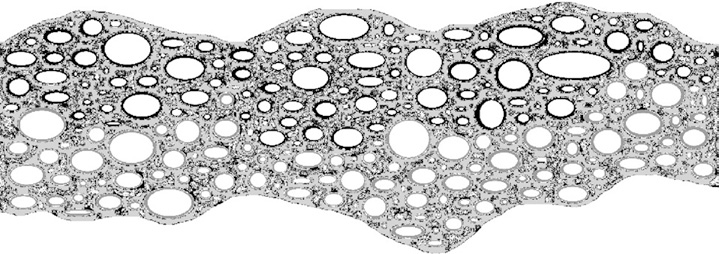
Рис. 3. Сухая фаза: минеральные и органические осадки из щелочной воды источников и соленой морской воды оставили отложения на стенках микроячеек (отмечены черным и серым цветом соответственно)

Рис. 4. Повторное заполнение во влажную фазу: граница раздела фаз проходит по новой линии, в результате чего часть ячеек с щелочными отложениями на стенках заполняется кислой средой и наоборот

Рис. 5. После нескольких циклов смены сухих и влажных фаз часть ячеек оказывается заполненными разнородными слоями, с возможным формированием биоэлектрического «вольтового столба» – аккумулятора энергии, а сами ячейки окружены ячейками с содержимым существенно иного рН

Рис. 6. Мембраны «научились» энергетически самоподдерживаться и самовосстанавливаться, возможно, с помощью структурных нуклеиновых кислот и рибозимов. Минеральная составляющая полупроницаемых мембран может впоследствии «вымываться», оставляя ассоциированные с новыми органическими оболочками каталитические железосерные кластеры
На близком принципе – чередования соленых и пресных фаз в изолированных элементах – уже построен ряд экспериментальных энергетических установок. С их помощью предполагается использовать «энтропийную» энергию градиента солености Мирового океана, оцениваемую до 1700 ТВт*ч/год (Skilhagen S.E., 2011), так называемую «голубую энергию» океана. Интересными прототипами можно назвать емкостную технологию итальянского инженера-физика Дориано Броджиоли (Doriano Borgioli, 2009) на основе двуслойного электролитического конденсатора (ионистора) большой емкости, работающего по сложному циклу заряда/разряда и поочередного наполнения камеры конденсатора соленой и пресной водой, и более продвинутую технологию группы исследователей из Стэнфордского университета (Ye M. et al., 2019), использующую «батареи энтропийного смешивания» (БЭС; Mixing Entropy Batteries, MEB). БЭС включает два электрода с большой поверхностью контакта, между которыми происходит чередование фаз морской и пресной воды. Один электрод состоит из берлинской лазури, смеси нескольких гексацианоферратов (II), весьма простых соединений железа, азота и углерода, другой представляет собой проводящий органический полимер, полипиррол. В циклическом процессе при заполнении камеры батареи пресной водой ионы натрия и хлориды выходят из соответствующих электродов в воду, создавая электрический ток между электродами. При смене пресной воды на морскую ионы натрия и хлориды забираются обратно в электроды, также образуя электрический ток, но уже обратной направленности.
Технически протонный градиент между средами с разным рН может быть заменен на натриевый и/или калиевый градиенты, имеющие бОльшую буферную емкость из-за большего содержания калия и натрия в рассматриваемых водах, нежели свободные протоны. На идее первичного калиевого градиента, наследуемого почти всеми формами жизни – преобладания калия внутри, а натрия снаружи клетки – основывается модель Армена Мулкиджаняна, Дарьи Дибровой, Михаила Гальперина и Евгения Кунина (Mulkijanian A. et al., 2012; Диброва Д. и соавт., 2015). В этой модели за основу для формирования протобиологических структур принимается конденсат испарений наземных геотермальных полей, в котором, как и в клеточных цитоплазмах, преобладает калий. Сам пресный конденсат неизбежно должен был находиться в щелочном окружении богатых натрием горячих геотермальных вод. При наличии адекватного полупроницаемого разделения на этой основе могла возникнуть первичная фосфатная, а затем самоподдерживающаяся натрий-калиевая мембранная энергетика. Как показывают недавние биоинформационные исследования группы Мулкиджаняна (Козлова М. И. и соавт., 2020), эта древнейшая натриевая энергетика («натриевый мир») не заместилась полностью более «современной» протонной энергетикой, а оказалась органично встроенной в конфигурацию биоэнергетических и биоинформационных процессов большинства современных эукариот, архей и бактерий. Даже у высших животных, включая человека, ключевые белки межклеточных коммуникаций – ассоциированные с G-белком рецепторы (GPCR, G-protein coupled receptors), одна из самых широко представленных в организме групп белков, – имеют прямое происхождение от древнейших белковых энергетических структур «натриевого мира».
Строительные элементы доклеточных структур
Было бы любопытно предположить, какие органические полимеры могли составить первичную органическую основу многослойных отложений на минеральных мембранах, обеспечив функциональный базис дальнейшего развития. Липиды и полисахариды, составляющие такую основу для большинства современных мембран живых организмов, возможны, но их синтез требует достаточно изощренных каталитических механизмов, практически невероятных на данном этапе развития биологической сложности. Белки также вполне могут подойти на эту роль, так как принципиально способны образовывать стабильные оболочки живых структур и способны к автокаталитическому самоподдержанию. Потенциальным недостатком может оказаться тот факт, что белки с (почти) случайной последовательностью аминокислот (в отсутствие еще носителей наследственной информации) чаще всего будут оказываться структурно неустойчивыми и неспособными к самоукладке в четвертичные (многомолекулярные) макрокомплексы. Для нуклеиновых кислот структурная функция не рассматривается в качестве первичной даже на ранних этапах эволюции. Обычно предполагается, что нуклеиновые кислоты эволюционно начинают как носители наследственной информации и катализаторы первых биохимических реакций. Однако и сейчас, в современных организмах, нуклеиновые кислоты зачастую выполняют структурную функцию даже у эволюционно далеких организмов, причем, и очень часто, в экстремальных для них условиях. При этом выполнение такой структурной функции нуклеиновыми кислотами не демонстрирует критической зависимости от четкой нуклеотидной последовательности. Так, нуклеиновые кислоты составляют основу гнойного содержимого ран – практически последней линии защиты при атаке патогенами физических повреждений многоклеточных организмов. Видимо, совсем неслучайно именно это содержимое стало местом открытия самих нуклеиновых кислот (ДНК) швейцарским врачом Иоганном Фридрихом Мишером в 1869 году.
Физиологической основой формирования воспалительных структур, основанных на ДНК – так называемых внеклеточных нитей нейтрофилов (NET, neutrophil extracellular traps), является процесс нетоза нейтрофилов. Нейтрофилы – наиболее распространенная разновидность лейкоцитов, рядовые солдаты острой воспалительной реакции организма. Сам нетоз можно считать разновидностью апоптоза – запрограммированной клеточной смерти, а нейтрофилы – клетками-камикадзе, предназначенными умереть, отдав самое святое – свою ДНК – в качестве кирпичей на строительство оборонительных рубежей всего организма (собственно смерть нейтрофила при этом не является совсем обязательной, а сам нетоз может создавать при определенных условиях и существенные проблемы для всего организма (Papayannopoulos V., 2018)). Важность этого защитного механизма, имеющего, вероятно, очень глубокие эволюционные корни – своего рода возврат к первичным основаниям жизни, своеобразное повторение филогенеза в патогенезе, подтверждается тем фактом, что наличие ферментов экзонуклеаз, способных разрушать NET структуры, служит существенным фактором вирулентности бактерий (Sharma P. et al., 2019). Напротив, и сами бактерии способны формировать из ДНК несущую решетку своего внеклеточного матрикса – основу бактериальных биопленок, своеобразный защитный чехол бактериального сообщества. Причем ключевые единицы такой ДНК-решетки аналогичны так называемым структурам Холидея – крестообразным формированиям двухцепочечной ДНК (рис. 7). И совершенно неслучайно именно ДНК-структуры, в том числе сформированные с использованием структур Холидея, применяются в качестве строительных блоков для ряда нанотехнологических материалов. Тем более что ДНК в качестве структурного материала обладает рядом уникальных свойств: например, высокоточной самосборкой и самоукладкой в полимолекулярные комплексы (Rothemund P. W. K., 2006), способностью проводить ток (в том числе фотохимического происхождения), что может быть весьма существенным фактором для участия в энергопроизводящих ячеистых структурах (см. рис. 2-6). Причем менее организованная, но более компактная А-форма (конформационно близкая к двухцепочечной РНК и ДНК-РНК гибридам) делает это на порядок более эффективно, чем более известная В-форма – правозакрученная спираль (Artes J. M. et al., 2015).
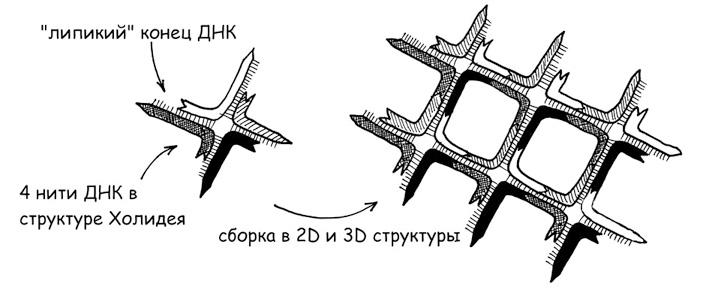
Рис. 7. Структуры Холидея и их искусственные производные
Рассматривается, что именно ДНК-РНК гибриды и могли быть первыми протобиологическими молекулами: Цзяньфен Сю и Джон Сазерленд с коллегами (Jianfeng Xu, John Sutherland et al., 2020) показали, что в кислой среде, в присутствии ионов натрия, магния, хлоридов и нитритов из простейших неорганических веществ типа цианидов может образовываться система нуклеозидов из 4 оснований, 2 из которых – пуриновые дезоксирибонуклеозиды (аденозин и инозин), и 2 пиримидиновые рибонуклеозиды (цитидин и уридин). До этого не удавалось продемонстрировать возможность возникновения рибозы и рибозидов в условиях первичной Земли (в отличие от дезоксирибозы и ее производных). В последнее время, при почти полном доминировании идеи РНК-мира (первичной роли РНК в возникновении жизни, ввиду ее способности как к сохранению информации, так и к катализу, в том числе способствующему саморепликации), появляются и другие свидетельства того, что РНК и ДНК, а возможно, и их гибриды, появились и могли сосуществовать еще до возникновения жизни (Extance A., 2020). В качестве одного из таких вариантов японскими исследователями из университета Нагоя (Murayama K. et al., 2021) предложены близкие к ДНК достаточно стабильные ксенонуклеиновые кислоты (КНК), в частности на основе L-треонинола (L-aТНК) – алифатической нуклеиновой кислоты (то есть без пуринового или пиримидинового колец, свойственных «нормальным» нуклеиновым кислотам). Особенностью полимеров данной кислоты, возможно, даже с включением «нормальных» нуклеотидов, является возможность самосборки (автополимеризации) в виде двойных цепей без участия белковых ферментов. Более того, фрагменты КНК способны далее сами выступать в качестве катализаторов самосборки, подобно некоторым РНК-фрагментам (рибозимам). Дополнительным свидетельством возможности сценария первичных структурных нуклеиновых кислот может служить обнаружение конъюгатов углеводов с некодирующими РНК – гликан-РНК – в составе мембран ряда организмов, где они, похоже, выполняют роль сигнальных рецепторов (Flynn R. A. et al., 2021). Этим самым открыт своеобразный мостик, возможно, очень древний, между миром генетической информации нуклеиновых кислот и миром разделительных и сигнальных углеводных структур.
Такие структуры возможны?
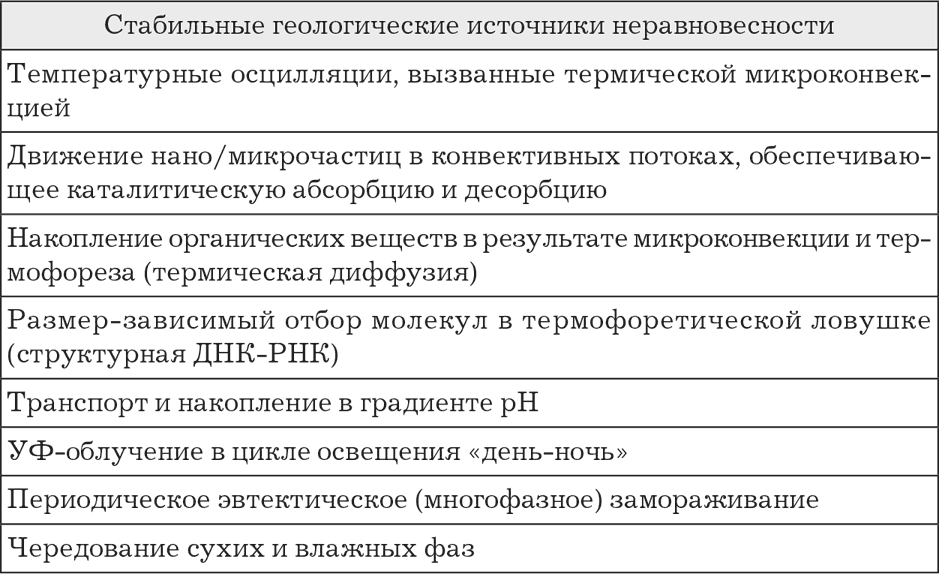
Важность разделительных структур в эволюции нуклеиновых кислот акцентируется в работах немецкого исследователя Кристофа Маста (Christof Mast et al., 2010; 2013). Мастом сформулирована теория термодинамической ловушки. Она показывает, что наличие термохимических градиентов создает дополнительные состояния неравновесности, стимулирующие в частных случаях полимеризацию, репликацию и концентрирование ДНК и возникновение дарвинского отбора в более общих случаях (табл. 1). Помимо термического градиента, вызванного геологическими условиями (разница температур гидротермального источника и внешней (например, морской воды), дополнительный, хотя и сравнительно микроскопический, но в некоторых ситуациях решающий вклад мог вносить и химический градиент протонов (электронов) и/или натрия, способный, в принципе, также создавать температурный градиент. Этот искусственно возникающий градиент, в свою очередь, несмотря на свою кажущуюся незначительность, мог способствовать впоследствии независимости протобиологических структур от геологических. По некоторым расчетам, даже незначительного температурного градиента в отдельных случаях может оказаться достаточно, например для обеспечения простого деления протоклеток: перемещение более «горячих» молекул двуслойной наружной мембраны наружу, движимое простыми физическими механизмами, увеличивает среднюю кривизну и усиливает любое локальное сжатие протоклетки вплоть до полного разделения на две (Romain Attal and Laurent Schwartz, 2021).
С другой стороны, мы видим, как уже на самых ранних этапах возникает двухвариантность реализации химического (протонного) градиента: 1) как концентрирование в форме химических связей (или в составе любых синтезируемых соединений, или, что оказывается выгоднее, в форме нескольких универсальных «энергетических валют», например АТФ); 2) как рассеивание (диссипация) энергии для более общей неспецифической модификации условий окружающей среды («контекста организации»).
Таблица 1. Примеры состояний геологической неравновесности (по Mast C. et al., 2010)
Насыщенные нуклеиновыми кислотами неорганические мембраны могли участвовать в трансмембранном и трансклеточном переносе энергии, что запустило бы эволюционный отбор нуклеиновых кислот. Впоследствии же возникший отбор нуклеиновых кислот с первоначально случайными последовательностями смог бы выявить ряд комбинаций, оказавшихся способных к саморепликации, и сделал «чистые» ДНК и РНК ключевыми операторами наследственной информации со своим четко определенным функционалом. В этом смысле первыми автореплицирующимися структурами могли быть общие предки вирусов и прокариот, паразитировавшие на длинных структурных/проводящих ДНК. Чередование кислотных и щелочных, влажных и сухих фаз в существовании доклеточных протобиологических структур можно сопоставить с чередованием хаотической и динамической фаз в теории эволюции информационных систем. Учитывая бóльшую чувствительность РНК к рН среды, щелочные влажные фазы могли выполнять роль хаотического «перемешивающего слоя» в протоэволюции нуклеиновых кислот и в большей степени могло быть связаны с РНК, а в кислотные или сухие фазы более стабильные ДНК фиксировали достижения и канализировали развитие (БОН: глава IV). Д. С. Чернавский, физик и математик, автор статистической теории «перемешивающих слоев», исходя из физических и информационных представлений, также считал ДНК более предпочтительной в качестве приоритетной молекулы при возникновении биологической сложности.
Есть основания предполагать, что эволюция протоживого и живого первые миллионы, если не миллиарды лет – от первых, еще не идентифицированных самореплицирующихся молекул до возникновения уцелевших до наших дней, хоть и в довольно измененном виде, архей и бактерий – проходила в значительной степени в привязке к горячим щелочным термальным источникам, подводным или наземным, являвшимися в первую очередь источниками химической энергии и минимальной упорядоченности, негэнтропии, которую и научились «есть» первые организмы.
В зависимости от того, с какой стороны кислотно-щелочного раздела им оказалось сподручнее усваивать эту негэнтропию, то есть в какую сторону оказался направлен протонный (протонно-натриевый?) насос – на выталкивание или запуск протонов в клетку – первые организмы разделились на линию бактерий (выталкивание протонов) и линию архей (запуск в клетку). Представляется, что самым ранним протобактериям и протоархеям энергетически выгоднее было бы сосуществование; само отнесение к протоархеям и протобактериям могло оказываться достаточно условным: направленность транспорта протонов в конкретной ячейке/протоклетке могла меняться в зависимости от изменения условий окружения в циклической геологической системе. Совершенствование биохимических механизмов в любом случае позволило им в конце концов разделиться и стать полностью самостоятельными. Уже самые древние независимые археи и бактерии имели между собой значительные различия как в ключевых наследственных генетических механизмах, так и структуре – в первую очередь в строении наружных оболочек на основе липидных полупроницаемых мембран. И когда этим двум сильно разошедшимся формам жизни вновь оказалось выгодным заново начать жить вместе в странном симбиозе, то, несмотря на громадные различия, накопившиеся за почти два миллиарда лет раздельного существования, позволившие успешно произойти этому событию только один раз и предопределившие все последующие проблемы совместного существования базовые энергетические механизмы археи и бактерии в новом симбиозе снова подошли друг к другу как ключ к замку. Бактерии, казалось бы, заняли в этом симбиозе подчиненное, «крепостное» положение маленьких органелл в большой архейной протоэукариотической клетке – с вроде бы как ограниченной ролью маленьких «электростанций», с невероятной эффективностью вырабатывающих огромное количество необходимой клетке энергии. Это сожительство дало начало как большей части наблюдаемой эволюции сложной жизни, так и ее предопределенной смерти.