



Weismann August
Studies in the Theory of Descent, Volume II
I should not have considered it worth while to have made these remarks if it had not been for the fact that the caterpillars of V. Io and V. Antiopa differ in small particulars from one another and from the other species of the genus. These differences relate to the number and position of the spines, as can be seen from the following table: —
Species of the Genus Vanessa, Fabr.
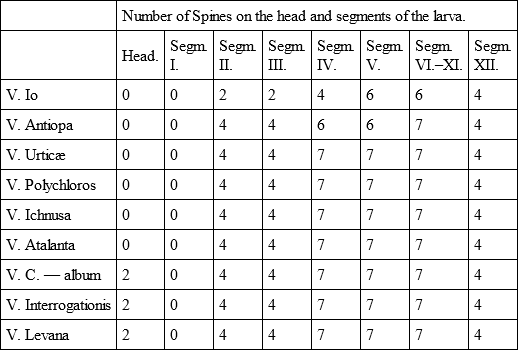
This character of the number of spines will not be considered as too unimportant when we observe how perfectly constant it remains in the nearly allied species. This is the case in the three consecutive forms, Urticæ, Polychloros, and Ichnusa. Now when we see that two species which differ in their imaginal characters present correspondingly small differences in their larvæ, this exact systematic congruence indicates a completely parallel phyletic development.
Exceptions are, however, to be met with here. Thus, Hübner has united one group of the species of Vanessa into the genus Pyrameis just mentioned, on account of certain characteristic distinctions of the butterflies. I do not know, however, how this genus admits of being grounded on the structure of the larvæ; the latter, as appears from the above table, agree exactly in the number and position of the spines with the caterpillars of Vanessa (sensû strictiori), nor can any common form of marking be detected which would enable them to be separated from Vanessa.
Still more striking is the incongruence in the genus Araschnia, Hübn. (A. Prorsa-Levana), which, like the genus Pyrameis, is entirely based on imaginal characters. This is distinguished from all the other sub-genera of the old genus Vanessa by a small difference in the venation of the wings (the discoidal cell of the hind-wings is open instead of closed). Now it is well-known that in butterflies the wing-venation, as most correctly shown by Herrich-Schäffer, is the safest criterion of “relationship.” It thus happens that this genus, typified by the common Levana, is in Kirby’s Catalogue separated from Vanessa by two genera, and according to Herrich-Schäffer18 by forty genera! Nevertheless, the larvæ agree so exactly in their spinal formula with Grapta that we should have no hesitation in regarding them as a species of this sub-genus. It appears to me very probable that in this case the form-relationship of the caterpillar gives more correct information as to the blood-relationship of the species than that of the imago – in any case the larvæ show a different form-relationship to the imagines.
Just as in the case of butterflies there are many genera of Sphingidæ which can be based on the structure of the larvæ, and which agree with those founded on the imagines.
Thus, the genus Macroglossa is characterized by a straight anal horn, a spherical head, and by a marking composed of longitudinal stripes, these characters not occurring elsewhere in this combination. The nearly allied genus Pterogon, on the other hand, cannot be based on the larvæ only, since not only is the marking of the adult larva very distinct in the different species, but the anal horn is present in two species, whilst in a third (P. Œnotheræ) it is replaced by a knob-like eye-spot. The genus Sphinx (sensû strictiori) is distinguished by the simple, curved caudal horn, the smooth, egg-shaped head and smooth skin, and by a marking mainly composed of seven oblique stripes. The genus Deilephila is distinguished from the preceding by a dorsal plate, situated on the prothorax and interrupting the marking, as well as by the pattern, which here consists of a subdorsal line with ring-spots more or less numerous and developed; the skin also is rough, “shagreened,” although it must be admitted that there are exceptions (Vespertilio). The genus Chærocampa admits also of being based on the form-relationship of its caterpillars, although this is certainly only possible by disregarding the marking and taking alone into consideration the peculiar pig-like form of the larvæ. The genus Acherontia, so nearly related to Sphinx, possesses in the doubly curved caudal horn a character common to the genus (three species known19). Finally may be mentioned the genus Smerinthus, of which the larvæ, by their anteriorly tapering form, their shagreened skin and almost triangular head with the apex upwards, their simply curved anal horn, and by their seven oblique stripes on each side, constitute a genus as sharply defined as that formed by the moths.
Although in all the systematic divisions hitherto treated of there are cases where the form-relationship of the larva does not completely coincide with that of the imago, such incongruences are of far more frequent occurrence in the smallest systematic group, viz. species.
The larvæ of two species have very frequently a much nearer form-relationship than their imagines. Thus, the caterpillars of Smerinthus Ocellatus and S. Populi are closely allied in structure, marking, and colouring, whilst the moths in these two last characters and in the form of the wings are widely separated.20 Judging from the larvæ we should expect to obtain two very similar moths, but in fact both Populi and Ocellatus have many near allies, and these closely related species sometimes possess larvæ which differ more widely than those of more distantly related species of imagines.
Thus, in Amur-land and North America there occur species of Smerinthus which closely resemble our Ocellatus in colour, marking, and form of wing, and which possess the characteristic large blue ocellus on the hind-wings. S. Excæcatus is quite correctly regarded as the representative American form of our Ocellatus, but its caterpillar, instead of being leaf-green, is of a chrome-yellow, and possesses dark green instead of white oblique stripes, and has moreover a number of red spots, and a red band on the head – in brief, in the very characters (colour and certain of the markings) in which the imagines completely agree it is widely different from Ocellatus. It appears also to be covered with short bristles, judging from Abbot and Smith’s figure.21
Just in the same way that the species having the nearest conceivable form-relationship to Ocellatus possesses a relatively strongly diverging larva, so does the nearest form-relation of Populi (imago) offer a parallel case. This species, which is also North American, lives on Juglans Alba. The imago of Smerinthus Juglandis differs considerably from S. Populi in the form of the wings, but it resembles the European species so closely in marking and colouring that no doubt can exist as to the near relationship of the two forms. The caterpillar of S. Juglandis,22 however, differs to a great extent from that of Populi in colour – it is not possible to confound these two larvæ; but those of Populi and Ocellatus are not only easily mistaken for one another, but are distinguished with difficulty even by experts.
In this same family of the Sphingidæ cases are not wanting in which, on the other hand, the moths are far more closely allied than the larvæ. This is especially striking in the genus Deilephila, eight species of which are allied in the imaginal state in a remarkable degree, whilst the larvæ differ greatly from one another in colour, and to as great an extent in marking. These eight species are D. Nicæa, Euphorbiæ, Dahlii, Galii, Livornica, Lineata, Zygophylli, and Hippophaës. Of these, Nicæa, Euphorbiæ, Dahlii, Zygophylli, and Hippophaës are so much alike in their whole structure, in the form of the wings, and in marking, that few entomologists can correctly identify them off-hand without comparison. The larvæ of these four species, however, are of very different appearances. Those of Euphorbiæ and Dahlii are most alike, both being distinguished by the possession of a double row of large ring-spots. Zygophylli (see Fig. 50, Pl. VI.) possesses only faint indications of ring-spots on a white subdorsal line; and in Hippophaës there is only an orange-red spot on the eleventh segment, the entire marking consisting of a subdorsal line on which, in some individuals, there are situated more or less developed ring-spots (see Figs. 59 and 60, Pl. VII.). If we only compare the larvæ and imagines of D. Euphorbiæ and Hippophaës, we cannot but be struck with astonishment at the great difference of form-relationship in the two stages of development.
In the case of D. Euphorbiæ and Nicæa this difference is almost greater. Whilst these larvæ show great differences in colour, marking, and in the roughness or smoothness of the skin (compare Fig. 51, Pl. VI. with Figs. 43 and 44, Pl. V.), the moths cannot be distinguished with certainty. As has already been stated, the imago of the rare D. Nicæa is for this reason wanting in most collections; it cannot be detected whether a specimen is genuine, i. e. whether it may not perhaps be a somewhat large example of D. Euphorbiæ.
An especially striking instance of incongruence is offered by the two species of Chærocampa most common with us, viz., Elpenor and Porcellus, the large and small Elephant Hawk-moths. The larvæ are so similar, even in the smallest details of marking, that they could scarcely be identified with certainty were it not that one species (Elpenor) is considerably larger and possesses a less curved caudal horn than the other. The moths of these two species much resemble one another in their dull green and red colours, but differ in the arrangement of these colours, i. e. in marking, and also in the form of their wings, to such an extent that Porcellus has been referred to the genus Pergesa23 of Walker. If systemy, as is admitted on many sides, has only to indicate the morphological relationship, this author is not to blame – but in this case a special larval classification must likewise be admitted, in a manner somewhat similar to that at present adopted provisionally in text-books of zoology for the Hydroid Polypes and inferior Medusæ. This case of Porcellus, however, shows that those are correct who maintain that systemy claims to express, although incompletely, the blood-relationship, and that systematists have always unconsciously formed their groups as though they intended to express the genetic connection of the forms. Only on this supposition can it appear incorrect to us to thus separate two species of which the larvæ agree so completely.
I cannot conclude this review of the various systematic groups without taking a glance at the groups comprised within species, viz. varieties. Whilst in species incongruence is of frequent occurrence, in varieties this is the rule, for which reason it admits in this case of being more sharply defined, since we are not concerned with a double difference but only with the question whether in the one stage a difference or an absolute similarity is observable. By far the majority of varieties are either simply imaginal or merely larval varieties – only the one stage diverges, the other is quite constant.
Thus, as has already been shown, in all the seasonally dimorphic butterflies known to me the caterpillars of the two generations of imagines, which are often so widely different, are exactly alike; and the same obtains for the majority of purely climatic varieties of butterflies. Unfortunately there are as yet no connected observations on this point. The only certain instance that I can here mention is that of the Alpine and Polar form of Pteris Napi. This variety, Bryoniæ, the female of which differs so greatly in marking and colouring, possesses larvæ which cannot be distinguished from those of the ordinary form of Napi.(See part I. appendix I. p. 124.)
That caterpillars can also vary locally without thereby affecting the imagines is shown by the frequently mentioned and closely investigated cases of di- and polymorphism in the larvæ of a number of Sphingidæ (M. Stellatarum, A. Atropos, S. Convolvuli, C. Elpenor, and Porcellus, &c.). The same thing is still more clearly shown by those instances in which there are not several but only one distinct larval form occurring in each of two different localities.
To this class belongs the above-mentioned case of Chærocampa Celerio (p. 197), supposing our information concerning this species to be correct; likewise the recently-mentioned case of the Ligurian variety of the caterpillar of Saturnia Carpini; and finally the case of Eriogaster Lanestris, so well known to lepidopterists. This insect inhabits the plains of Germany, and in the Alps extends to an elevation of 7000 feet, where it possesses a larva differently marked and coloured (E. Arbusculæ) to those of the lowlands whilst the moths are smaller, but do not differ in other respects from those of the plains.
Among the Alpine species many other such cases may occur, but these could only be discovered by making investigations having special reference to this point. Of the Alpine butterflies, for example, not a single species can have been reared from the caterpillar; for this reason but few observations have on the whole been given by entomologists respecting the Alpine larvæ, which are not known sufficiently well to enable such a question to be decided.
The investigation of the form-relationships existing between larvæ on the one hand and imagines on the other has thus led to the following results: —
We learn on comparison that incongruences or inequalities of form-relationship occur in all systematic groups from varieties to families. These incongruences are of two kinds, in some cases being disclosed by the fact that the larvæ of two systematic groups, e. g. two species, are more closely related in form than their imagines (or inversely), whilst in other cases the larvæ form different systematic groups to those formed by the imagines.
The results of the investigation into the occurrence of incongruences among the various systematic groups may be thus briefly summarised: —
Incongruences appear to occur most frequently among varieties, since it very frequently happens that it is only the larva or only the imago which has diverged into a variety, the other stage remaining monomorphic. The systematic division of varieties is thus very often one-sided.
Among species also incongruences are of frequent occurrence. Sometimes the imagines are much more nearly related in form than the larvæ, and at others the reverse happens; whilst again the case appears also to occur in which only the one stage (larva) diverges to the extent of specific difference, the other stage remaining monomorphic (D. Euphorbiæ and Nicæa).
The agreement in form-relationship appears to be most complete in genera. In the greater number of cases the larval and imaginal genera coincide, not only in the sharpness of their limits, but also – as far as one can judge – in the weight of their distinctive characters, and therefore in the amount of their divergence. Of all the systematic groups, genera show the greatest congruence.
In families there is again an increase of irregularity. Although larval and imaginal families generally agree, there are so many exceptions that the groups would be smaller if they were based exclusively on the larval structure than if founded entirely on the imagines (Nymphalidæ, Bombycidæ).
If we turn to the groups of families we find a considerably increased incongruence; complete agreement is here again rather the exception, and it further happens in these cases that it is always the larvæ which, to a certain extent, remain at a lower grade, and which form well defined families; but these can seldom be associated into groups of a higher order having a common character, as in the case of the imagines (Rhopalocera).
After having thus collected (so far as I am able) the facts, we have now to attempt their interpretation, and from the observed congruence and incongruence of form-relationship of the two stages to endeavour to draw a conclusion as to the underlying causes of the transformations.
It is clear at starting that all cases of incongruence can only be the expression or the consequence of a phyletic development which has not been exactly parallel in the two stages of larva and imago – that one stage must have changed either more rapidly or more slowly than the other. An “unequal phyletic development” is thus the immediate cause of incongruence.
Thus, the occurrence of different larvæ in species of which the imagines have remained alike may be simply understood as cases in which the imago only has experienced a change – has taken a forward step in phyletic development, whilst the larvæ have remained behind. If we conceive this one-sided development to be repeated several times, there would arise two larval forms as widely different as those of Deilephila Nicæa, and Euphorbiæ, whilst the imagines, as is actually the case in these species, would remain the same.
The more commonly occurring case in which one stage has a greater form-divergence than the other, is explicable by the one stage having changed more frequently or more strongly than the other.
The explanation of the phenomena thus far lies on the surface, and it is scarcely possible to advance any other; but why should one stage become changed more frequently or to a greater extent than the other? why should one portion be induced to change more frequently or more strongly than another? whence come these inducements to change? These questions bring us to the main point of inquiry: – Are the causes which give rise to these changes internal or external? Are the latter the result of a phyletic vital force, or are they only due to the action of the external conditions of life?
Although an answer to this question will be found in the preceding essay, I will not support myself on the results there obtained, but will endeavour to give another solution of the problem on fresh grounds. The answer will indeed be the same as before: – A phyletic force must be discountenanced, since in the first place it does not explain the phenomena, and in the second place the phenomena can be well explained without its assumption.
The admission of a phyletic vital force does not explain the phenomena. The assumption that there is a transforming power innate in the organism indeed agrees quite well with the phenomenon of congruence, but not with that of incongruence. Since a large number of cases of the latter depend upon the fact that the larvæ are more frequently influenced by causes of change than their imagines, or vice versâ, how can this be reconciled with such an internal force? On this assumption would not each stage of a species be compelled to change, if not contemporaneously at least successively, with the same frequency and intensity, by the action of an innate force? and how by means of the latter can there ever result a greater form-divergence in the larvæ than in the imagines?
It is delusive to believe that these unequal deviations can be explained by assuming that the phyletic force acts periodically. Granting that it does so, and that the internal power successively compels the imago, pupa, and finally the larva to change, there would then pass a kind of wave of transformation over the different stages of the species, as was actually shown above to be the case in the single larval stages. The only possible way of explaining the unequal distances between larvæ and imagines would therefore be to assume that two allied groups, e. g. species, were not contemporaneously affected by the wave, so that at a certain period of time the imago alone of one species had become changed, whilst in the other species the wave of transformation had also reached the larva. In this case the imagines of the two species would thus appear to be more nearly related than their larvæ.
Now this strained explanation is eminently inapplicable to varieties, still less to species, and least of all to higher systematic groups, for the simple reason that every wave of transformation may be assumed to be at the most of such strength as to produce a deviation of form equal to that of a variety. Were the change resulting from a single disturbance greater, we should not only find one-sided varieties, i. e. those belonging to one stage, but we should also meet as frequently with one-sided species. If, however, a wave of transformation can only produce a variety even in the case of greatest form-divergence, the above hypothetical uncontemporaneous action of such a wave in two species could only give rise to such small differences in the two stages that we could but designate them as varieties. An accumulation of the results of the action of several successive waves passing over the same species could not happen, because the distance from a neighbouring species would always become the same in two stages as soon as one wave had ended its course. In this manner there could therefore only arise divergences of the value of varieties, and incongruences in systematic groups of a higher rank could not thus be explained.
All explanations of the second form of incongruence from the point of view of a phyletic force can also be shown to be absurd. How can the fact be explained that larval and imaginal families by no means always coincide; or that the larvæ can only be formed into families whilst the imagines partly form sharply defined groups of a higher order? How can an internal directive force within the same organism urge in two quite distinct directions? If the evolution of a definite system were designed, and the admission of such a continually acting power rendered necessary, why such an incomplete, uncertain, and confused performance?
I must leave others to answer these questions; to me a vital force appears to be inadmissible, not only because we cannot understand the phenomena by its aid, but above all because it is superfluous for their explanation. In accordance with general principles the assumption of an unknown force can, however, only be made when it is indispensable to the comprehension of the phenomena.
I believe that the phenomena can be quite well understood without any such assumption – both the phenomena of congruence and incongruence, in their two forms of unequal divergence and unequal group-formation.
Let us in the first place admit that there is no directive force in the organism inciting periodic change, but that every change is always the consequence of external conditions, being ultimately nothing but the reaction – the response of the organism to some of the influences proceeding from the environment; every living form would in this case remain constant so long as it was not compelled to change by inciting causes. Such transforming factors can act directly or indirectly, i. e. they can produce new changes immediately, or can bring about a remodelling by the combination, accumulation, or suppression of individual variations already present (adaptation by natural selection). Both forms of this action of external influences have long been shown to be in actual operation, so that no new assumption will be made, but only an attempt to explain the phenomena in question by the sole action of these known factors of species formation.
If, in the first instance, we fix our attention upon that form of incongruence which manifests itself through unequal divergence of form-relationship, it will appear prominently that this bears precise relations to the different systematic groups. This form of incongruence constitutes the rule in varieties of the order Lepidoptera, it is of very frequent occurrence in species, but disappears almost completely in genera, and entirely in the case of families and the higher groups. On the whole, therefore, as we turn to more and more comprehensive groups, the incongruence diminishes whilst the congruence increases, until finally the latter becomes the rule.
Now if congruence presupposes an equal number of transforming impulses, we perceive that the number of the impulses which have affected larvæ and imagines agree with one another the more closely the larger the systematic groups which are compared together. How can this be otherwise? The larger the systematic group the longer the period of time which must have been necessary for its formation, and the more numerous the transforming impulses which must have acted upon it before its formation was completed.
But if the supposition that the impulse to change always comes from the environment in no way favours the idea that such impulses always affect both stages contemporaneously, and are equal in number during the same period of time, there is not, on the other hand, the least ground for assuming that throughout long periods the larvæ or the imagines only would have been affected by such transforming influences. This could have been inferred from the fact that varieties frequently depend only upon one stage, whilst specific differences in larvæ only also occur occasionally, the imagines remaining alike; but no single genus is known of which all the species possess similar larvæ. Within the period of time during which genera can be formed the transforming impulses therefore never actually affect the one stage only, but always influence both.
But if this is the case – if within the period of time which is sufficient for the production of species, the one stage only is but seldom and quite exceptionally influenced by transforming impulses, whilst both stages are as a rule affected, although not with the same frequency, it must necessarily follow that on the whole, as the period of time increases, the difference in the number of these impulses which affect the larva and of those which affect the imago must continually decrease, and with this difference the magnitude of the morphological differences resulting from the transforming influences must at the same time also diminish. With the number of the successively increasing changes the difference in the magnitude of the change in the two stages would always relatively diminish until it had quite vanished from our perception; just in the same manner as we can distinguish a group of three grains of corn from one composed of six, but not a heap of 103 grains from one containing 106 grains.
That the small systematic groups must have required a short period and the large groups a long period of time for their formation requires no special proof, but results immediately from the theory of descent.
All the foregoing considerations would, however, only hold good if the transforming impulses were equal in strength, or, not to speak figuratively, if the changes only occurred in equivalent portions of the body, i. e. in such portions as those in which the changes are of the same physiological and morphological importance to the whole organism.
Now in the lower systematic groups this is always the case. Varieties, species, and genera are always distinguished by only relatively small differences; deep-seated distinctions do not here occur, as is implied in the conception of these categories. The true cause of this is, I believe, to be found in the circumstance that all changes take place only by the smallest steps, so that greater differences can only arise in the course of longer periods of time, within which a great number of types (species) can, however, come into existence, and these would be related by blood and in form in different degrees, and would therefore form a systematic group of a higher rank.
The short periods necessary for the production of inferior groups, such as genera, would not result in incongruences if only untypical parts of the larvæ, such as marking or spines, underwent change, whilst in the imagines typical parts – wings and legs – became transformed. The changes which could have occurred in the wings, &c., during this period of time would have been much too small to produce any considerable influence on the other parts of the body by correlation; and two species of which the larvæ and imagines, had changed with the same frequency, would show a similar amount of divergence between the larvæ and between the imagines, although on the one side only untypical parts —i. e. those of no importance to the whole organization – and on the other side typical parts, were affected. The number of the changes would here alone determine whether congruence or incongruence occurred between the two stages.
The case would be quite different if, throughout a long period of time, in the one stage only typical and in the other only untypical parts were subjected to change. In the first case a complete transformation of the whole structure would occur, since not only would the typical parts, such as the wings, undergo a much further and increasing transformation in the same direction, but these changes would also lead to secondary alterations.
In this manner, I believe, must be explained the fact that in the higher groups still greater form-divergences of the two stages occur; and if this explanation is correct, the cause of this striking phenomenon, viz., that incongruence diminishes from varieties to genera, in which latter it occurs but exceptionally, whilst in families and in the higher groups it again continually increases, is likewise revealed. Up to genera the incongruence depends entirely upon the one stage having become changed more frequently than the other; but in families and groups of families, and in the orders Diptera and Hymenoptera, as will be shown subsequently, in sub-orders and tribes, it depends upon the importance of the part of the body affected by the predominant change. In the latter case the number of changes is of no importance, because these are so numerous that the difference vanishes from our perception; but an equal number of changes, even when very great, may now produce a much greater or a much smaller transformation in the entire bodily structure according as they affect typical or untypical portions, or according as they keep in the same direction throughout a long period of time, or change their direction frequently.
Those unequal form-divergences which occur in the higher systematic groups a re always associated with a different formation of groups – the larvæ form different systematic groups to the imagines, so that one of these stages constitutes a higher or a lower group; or else the groups are of equal importance in the two stages, but are of unequal magnitude – they do not coincide, but the one overlaps the other.


