



Сергей Бабичев
Медицинская микробиология, иммунология и вирусология
Способы питания
Углеродное питание
К числу важнейших химических элементов-органогенов, необходимых для синтеза органических соединений, относят: углерод, азот, водород и кислород. Свою потребность в водороде и кислороде бактерии удовлетворяют за счет воды. Сложнее обстоит дело с углеродным и азотным питанием. По способу углеродного питания бактерии делят на аутотрофы и гетеротрофы.
Аутотрофы (греч. autos – сам, trophe – питание) – организмы, которые полностью удовлетворяют свои потребности в углероде за счет CO2.
Гетеротрофы (греч. heteros – другой, trophe – питание, т. е. «питаемые другими») – организмы, которые не могут удовлетворить свои потребности в углероде только за счет CO2, а требуют для питания готовых органических соединений. В свою очередь, гетеротрофов подразделяют на сапрофитов (греч. sapros – гнилой, phyton – растение), т. е. гетеротрофов, источником питания которых служат мертвые органические субстраты; и паразитов (греч. para – при, sitos – пища), т. е. гетеротрофов, живущих за счет живых тканей животных и растений. Для превращения CO2 в органические соединения требуется энергия: чтобы восстановить CO2 в один моль гексозы требуется около 112 ккал. Существует два источника этой энергии – фотосинтез и хемосинтез.
ФОТОСИНТЕЗ
Фотосинтез – это синтез за счет энергии солнечного света: свободная энергия фотона красного света (680 нм) ΔG = 41 ккал/моль, голубого (400 нм) – ΔG = = 65 ккал/моль. В результате фотосинтеза растительность земного шара ежегодно синтезирует более 100 млрд тонн органических веществ. На это используется около 200 млрд тонн CO2, и в атмосферу выделяется около 145 млрд тонн свободного O2. Общее количество солнечной энергии, используемой ежегодно для фотосинтеза, составляет не менее 3 ⋅ 1021 Дж.
У растений мобилизация солнечной энергии и превращение ее в энергию химических связей, главным образом в виде АТФ, осуществляется с помощью хлоропластов, содержащих хлорофилл (греч. chloros – зеленый, phyllon – лист) – зеленый пигмент, связанный с белками и липидами их мембран. Основу молекулы хлорофилла составляет магниевый комплекс порфиринового цикла, близкий к другим комплексам порфирина (с железом) – цитохромам, гему и т. п. Поглощая энергию фотона солнечного света, электрон в молекуле хлорофилла возбуждается и переходит с основного энергетического уровня на более высокий, а затем он стремится вновь возвратиться на свой основной, стабильный энергетический уровень, отдавая при этом поглощенную энергию. Если такое возвращение происходит в чистом препарате хлорофилла, поглощенная энергия испускается в виде света. Однако в клетке хлорофилл связан со специфическими молекулами, и поэтому возбужденный электрон отрывается от молекулы хлорофилла и транспортируется этими молекулами – переносчиками электронов. Они передают его по замкнутой цепи реакции вне молекулы хлорофилла. Возбужденный электрон постепенно отдает свою энергию, которая и используется для синтеза АТФ из АДФ и неорганического фосфора, а далее электрон возвращается на свое прежнее место в молекуле хлорофилла, способной после этого поглощать другой фотон. В ходе переноса возбужденного электрона по крайней мере два из его переносчиков способны мобилизовать часть переносимой им энергии для синтеза АТФ (рис. 18). В процессе фотосинтеза происходит не только связывание солнечной энергии, но и синтез углеводов, в частности глюкозы, путем восстановления CO2, т. е. добавления к ней электронов и водорода. Источником электронов служит хлорофилл, а источником протонов – вода.
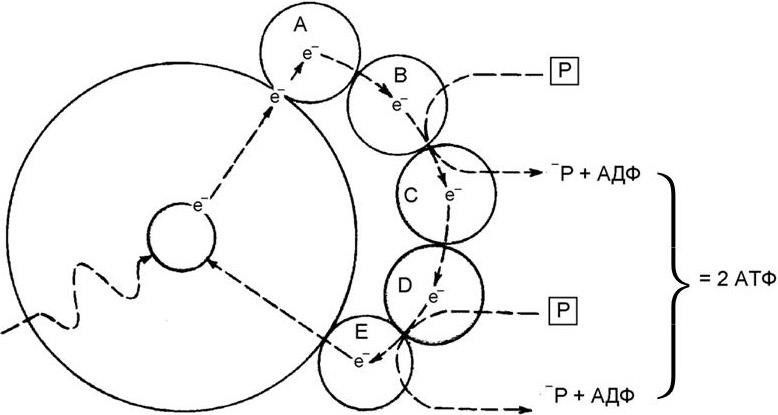
Рис. 18. Процесс циклического фосфорилирования, посредством которого в молекуле хлорофилла электрон, перенесенный на более высокий энергетический уровень благодаря поглощению фотона света, обеспечивает энергию, необходимую для образования АТФ (по А. Ленингеру)
Реакция фотосинтеза осуществляется в две фазы. В первой (световые реакции) – под действием фотонов электрон хлорофилла переходит в возбужденное состояние;
затем, возвращаясь в свое обычное энергетическое состояние, он освобождает энергию, которая используется для синтеза таких молекул, как АТФ и НАДФН2. Во второй фазе (темновые реакции) АТФ и НАДФН2 используются для восстановления CO2 в глюкозу.
Суммарная реакция фотосинтеза такова:

Таким образом, молекула глюкозы, которая представляет собой конечный продукт фотосинтеза (наряду с кислородом), содержит большое количество солнечной энергии (около 690 ккал на 1 моль), заключенной в ее молекулярной структуре. Гетеротрофные организмы извлекают эту энергию путем последовательного расщепления молекулы глюкозы для того, чтобы «законсервировать» содержащуюся энергию в форме вновь образующихся молекул АТФ – универсальных хранителей и носителей энергии, необходимой для жизни всех клеток.
К фотосинтезирующим бактериям – фототрофам – относятся цианобактерии (сине-зеленые водоросли), пурпурные и зеленые бактерии, а также некоторые архебактерии.
Цианобактерии – различные многоклеточные нитчатые и одноклеточные организмы, среди них есть подвижные формы, которые передвигаются с помощью скольжения. У цианобактерий, как и у растений, фотосинтез осуществляется с помощью хлорофилла и сопровождается выделением свободного кислорода.
Архебактерии (экстремальные галофилы) осуществляют особую форму фотосинтеза. У них вместо хлорофилла в фотосинтезе участвует особый пигмент – бактериородопсин (комплекс каротиноида ретиналя с белком), который под влиянием света претерпевает фотохимические превращения, непосредственно сопряженные с синтезом АТФ.
Пурпурные и зеленые бактерии содержат различные по составу хлорофиллы (бактериохлорофиллы a, b, c, d и e) и каротиноиды. Большинство зеленых бактерий – мелкие, неподвижные грамотрицательные палочки. Пурпурные бактерии представлены грамотрицательными палочками, кокками или спириллами и часто имеют жгутики. У пурпурных бактерий хлорофилл замаскирован пурпурно-красным или коричневым пигментом, а фотосинтезирующий аппарат заключен в клеточную мембрану, у зеленых – в клеточную мембрану или в специальные органеллы – хлоробиум-везикулы, локализованные в цитоплазме или мембране. В отличие от растений, водорослей и цианобактерий при фотосинтезе пурпурные и зеленые бактерии не выделяют O2, так как для восстановления CO2 они используют в качестве доноров электронов не водород H2O, а водород H2S. При этом пурпурные бактерии окисляют H2S до H2SO4:
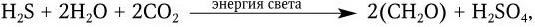
а зеленые серобактерии – до S2:

Некоторые пурпурные и зеленые бактерии в качестве донора электронов используют серу и другие ее неорганические соединения (тиосульфат, сульфит). Все они являются обитателями водоемов, где имеются наиболее благоприятные для них сочетания анаэробных условий, света и источников питания.
ХЕМОСИНТЕЗ
Русским ученым С. Н. Виноградским в 1880 – 1890 гг. было обнаружено, что некоторые грамотрицательные бактерии используют для своего роста энергию хемосинтеза, т. е. энергию, получаемую за счет окисления неорганических соединений. Такие организмы получили название хемоавтотрофов. Им было установлено, что хемоавтотрофы характеризуются двумя важнейшими особенностями:
1. Обладают высокой специфичностью в отношении неорганического источника энергии.
2. Очень часто они не только не способны использовать в качестве источников углерода и энергии органические вещества, но последние даже могут угнетать их рост.
К хемоавтотрофам относятся открытые С. Н. Виноградским нитрифицирующие бактерии, представленные двумя группами. Представители одной из них (роды Nitrosomonas, Nitrosospira, Nitrosococcus, Nitrosoglobus) окисляют NH3 до азотистой кислоты:

Представители другой (роды Nitrobacter, Nitrospira, Nitrococcus) окисляют азотистую кислоту до азотной:

Выделяемая при хемосинтезе энергия используется нитрифицирующими бактериями для ассимиляции CO2 и восстановления ее до глюкозы или других углеводов. Наиболее многочисленную и разнообразную группу хемосинтезирующих бактерий составляют водородные бактерии, осуществляющие реакцию:
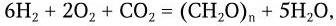
где (CH2O)n – условное обозначение синтезируемого углевода.
Но эти бактерии являются факультативными хемоавтотрофами, так как способны расти на средах, содержащих органические вещества. Некоторые строго анаэробные метанообразующие бактерии осуществляют хемосинтез по реакции:
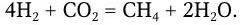
К хемоавтотрофам, как показал С. Н. Виноградский, относятся нитчатые скользящие бактерии Beggiatoa, Thiothrix и другие аэробы, способные окислять сероводород и тиосульфат до серы и сульфата. Внутри таких клеток часто обнаруживается сера.
К числу хемоавтотрофов относятся и некоторые виды железобактерий, в частности рода Gallionella, которые в качестве минерального источника восстановленного железа используют осадок сульфида железа.
У всех хемоавтотрофов ассимиляция CO2 происходит с помощью реакции, катализируемой рибулозодифосфаткарбоксилазой (реакция Кальвина):

Первичным продуктом фиксации CO2 является 3-фосфоглицериновая кислота, из которой синтезируются все остальные органические молекулы клетки.
Для осуществления ассимиляции CO2 хемоавтотрофы путем окислительной диссимиляции неорганического субстрата получают АТФ и восстановитель (восстановленный пиридиннуклеотид). Нитрифицирующие бактерии и многие тиобациллы имеют особые характерные для прокариот образования – карбоксисомы, содержащие рибулозодифосфаткарбоксилазу.
В зависимости от того, какими донорами электронов пользуются бактерии, их разделяют на литотрофы (используют неорганические доноры электронов H2, NH3, H2S, Fe и др.) и органотрофы (в качестве доноров электронов используют органические соединения).
Таким образом, по способу углеродного питания все организмы можно подразделить на три основные группы:
1. Фотолитотрофы (источник энергии – солнечный свет, доноры электронов – неорганические соединения).
2. Хемолитотрофы (источник энергии – окислительно-восстановительные реакции, доноры электронов – неорганические соединения).
3. Хемоорганотрофы (источник энергии – окислительно-восстановительные реакции, доноры электронов – органические соединения). Большинство патогенных бактерий относится к хемоорганотрофам.
Азотное питание
По способу азотного питания бактерии подразделяют на аминоавтотрофов и аминогетеротрофов. Аминоавтотрофы способны полностью удовлетворять свои потребности в азоте, необходимом для синтеза главным образом белков и нуклеиновых кислот, с помощью атмосферного или минерального азота. Аминогетеротрофы нуждаются для своего питания в готовых органических азотистых соединениях: некоторых аминокислотах, основаниях, витаминах и др.
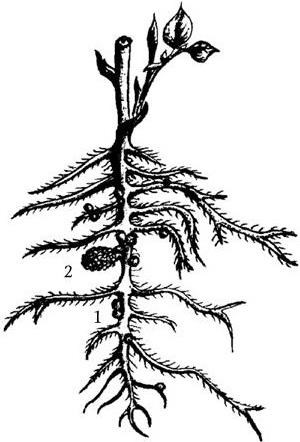
Рис. 19. Рисунок Мальпиги (XVII в.), изображающий корни бобового растения с корневыми клубеньками (1) и оболочкой семени (2), из которого развилось данное растение
К числу аминоавтотрофов относятся азотфиксирующие бактерии, свободно живущие в почве, и так называемые клубеньковые азотфиксирующие бактерии. Первый представитель азотфиксирующих бактерий – Clostridium pasteurianum (анаэроб) – был открыт в 1893 г. С. Н. Виноградским. В 1901 г. М. Бейеринк установил, что способностью усваивать атмосферный азот обладают также аэробные бактерии Azotobacter, занимающие по азотфиксирующей активности первое место (до 25 г азота на 1 кг использованного сахара), но менее распространенные в почве, чем C. pasteurianum. Азотфиксирующими свойствами обладают некоторые виды Pseudomonas, Bacillus, цианобактерий, а также пурпурные и зеленые серобактерии. Азотфиксирующие бактерии рода
Rhizobium получили название клубеньковых потому, что они, размножаясь на корнях ряда бобовых растений, образуют выросты-клубеньки (рис. 19). Размножаясь в них, они превращаются из мелких палочек в разветвленные формы – бактероиды, которые наиболее интенсивно связывают молекулярный азот. Симбиоз клубеньковых бактерий с растениями взаимовыгоден, так как Rhizobium продуцируют ряд физиологически активных соединений, которые благоприятно влияют на бобовые растения. После разрушения клубеньков клубеньковые бактерии живут в почве как сапрофиты.
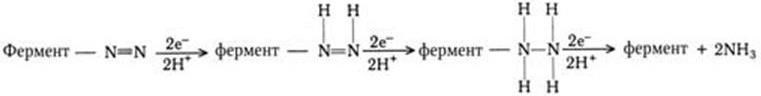
Азотфиксирующие бактерии восстанавливают азот (N2) до NH3 с помощью сложной ферментной системы нитрогеназы, содержащей железо, молибден, магний. Эта система нуждается в источнике электронов, которые поступают через восстановитель с низким потенциалом, содержащий негеминовое железо – ферредоксин (переносчик электронов).

Цепь переноса электронов состоит из ферредоксина (Фд), азоферредоксина (АзоФд) и молибдоферредоксина (МоФд), и за один раз переносятся только два электрона. Для последнего переноса расходуется 1 молекула АТФ.
Поскольку для восстановления N2 до NH3 требуется шесть электронов, реакция, очевидно, протекает через три последовательные двухэлектронные стадии:
Подробное выяснение механизма генетического контроля азотфиксирующих систем бактерий может создать необходимые предпосылки для искусственного переноса оперонов этих систем в геном растений и конструирования трансгенных растительных организмов, обладающих азотфиксирующей активностью, что имело бы огромное значение для сельского хозяйства.
Вторая большая группа аминоавтотрофов представлена нитрифицирующими бактериями, которые используют для синтеза белков в качестве основных источников азота соли аммиака, азотистой и азотной кислот. Подсчитано, что на образование вновь вырастающих растений ежегодно требуется около 1,5 млрд тонн азота в форме, доступной растениям. Поэтому нельзя не отметить, что азотфиксирующим и нитрифицирующим бактериям принадлежит исключительно важная роль в обеспечении плодородия почвы (см. раздел «Роль микроорганизмов в круговороте веществ в природе», гл. 15).
Аминогетеротрофы для роста и размножения нуждаются в различных органических азотистых соединениях. Необходимо оговориться, что аминогетеротрофы представляют собой также неоднородную группу, так как многие из них нуждаются для роста лишь в определенных аминокислотах, как, например, Francisella tularensis. Для ее роста требуется добавление к среде по крайней мере 10 аминокислот. В то же время многие бактерии, синтезируя аминокислоты и основания из минеральных источников азота, нуждаются в тех витаминах (ростовых факторах), которые сами не способны синтезировать: биотин (Н), тиамин (В1), рибофлавин (В2), пантотеновая кислота (В3), холин (В4), никотинамид (В5), фолиевая кислота (В9) и ее производные (В11) и т. п. Наконец, есть патогенные бактерии, например группа гемоглобинофильных организмов (Haemophilus), которые для роста нуждаются в добавлении к питательной среде сложных веществ, содержащихся в крови: Х-факторов (гемин) и др. Кроме того, в результате различных мутаций аминогетеротрофные бактерии могут превращаться в мутанты, неспособные синтезировать тот или иной метаболит и поэтому нуждающиеся в нем. Такие мутанты получили название ауксотрофов (лат. auxilium – помощь и греч. trophe – питание). Они во многом способствовали изучению особенностей биохимии бактерий.
Для нормальной жизнедеятельности бактерии, как и другие организмы, обязательно нуждаются в ионах Na+, K+, Cl-, Ca2+, Mg2+, Mn2+, Fe2+, Cu2+, а также в фосфоре и сере, которые поступают в клетку путем диффузии и активного транспорта.
Все процессы обмена веществ представляют собой цепь взаимосвязанных во времени и пространстве саморегулируемых реакций. Каждая из них и их совокупные пути катализируются соответствующими ферментами.
Ферменты
Ферменты (греч. fermentum – закваска), или энзимы, – специфические белковые катализаторы, присутствующие во всех живых клетках. Их нет у плазмид, у некоторых вирусов. Ферменты снижают энергию активации, которая необходима для осуществления той или иной химической реакции. Они направляют ее обходным путем через промежуточные реакции, требующие значительно меньшей энергии активации. Под влиянием ферментов происходит перераспределение электронных плотностей и некоторая деформация молекул субстрата, наступающая при образовании промежуточного фермент-субстратного комплекса. Эта деформация приводит к ослаблению внутримолекулярных связей и, следовательно, к понижению необходимой энергии активации, в результате чего скорость реакции возрастает. Открытию различных ферментов и изучению путей биохимических реакций, катализируемых ими, во многом способствовали работы с использованием в качестве объектов исследования бактерий, особенно ауксотрофов. У бактерий обнаружены все шесть классов ферментов:
1) оксидоредуктазы (катализируют окислительно-восстановительные реакции);
2) трансферазы (катализируют реакции переноса групп атомов);
3) гидролазы (катализируют гидролитическое расщепление различных соединений);
4) лиазы (катализируют реакции отщепления от субстрата той или иной химической группы негидролитическими путями с образованием двойной связи или, наоборот, присоединение химической группы к двойным связям);
5) изомеразы (катализируют внутримолекулярные превращения);
6) лигазы, или синтетазы (катализируют соединение двух молекул, сопряженное с расщеплением пирофосфатной связи в молекуле АТФ или аналогичного трифосфата).
Изучение ферментов, обнаруживаемых у бактерий, представляет большой интерес для микробиологической промышленности. Изучение особенностей обмена веществ патогенных бактерий необходимо прежде всего для понимания механизмов, с помощью которых они реализуют свою патогенность, т. е. для выяснения сущности патогенеза инфекционных заболеваний. Изучение биохимических свойств бактерий широко используется как для их систематики и классификации, так и для идентификации, т. е. для диагностики.
У бактерий обнаружены уникальные генетические механизмы контроля биосинтеза ферментов, они проявляются в виде феноменов индукции и репрессии. Индукция заключается в том, что синтез ферментов наступает только в присутствии специфических химических веществ, которые являются субстратом для данного фермента или аналогом этого субстрата. Например, синтез ферментов, участвующих в потреблении лактозы у E. coli, начинается (индуцируется) и происходит только при наличии в среде лактозы. Как только она исчезает, синтез этих ферментов прекращается.
Под эффектом репрессии понимают явление, при котором синтез фермента подавляется (репрессируется) под влиянием специфических химических соединений, почти всегда являющихся непосредственными продуктами (или аналогами продуктов) реакции, катализируемой этим ферментом. Например, синтез ферментов, участвующих в образовании метионина у E. coli, прекращается, как только в среде накапливается избыток этой аминокислоты. Нетрудно видеть, насколько совершенен такой механизм саморегуляции биохимических процессов.
В соответствии с этими особенностями генетического контроля, у бактерий различают три основные группы ферментов: конститутивные, синтез которых происходит в течение всего клеточного цикла; индуцибельные, синтез которых индуцируется соответствующим субстратом; и репрессибельные, синтез которых подавляется в результате избыточного накопления продукта реакции, катализируемой данным ферментом (ферментами).
Метаболизм
Биохимические процессы, протекающие в клетке, объединены одним словом – метаболизм (греч. metabole – превращение). Этот термин равнозначен понятию «обмен веществ и энергии». Различают две стороны метаболизма: анаболизм и катаболизм.
Анаболизм – совокупность биохимических реакций, осуществляющих синтез компонентов клетки, т. е. та сторона обмена веществ, которую называют конструктивным обменом.
Катаболизм – совокупность реакций, обеспечивающих клетку энергией, необходимой, в частности, и для реакций конструктивного обмена. Поэтому катаболизм определяют еще как энергетический обмен клетки.
В конструктивном обмене можно выделить две группы биосинтетических процессов: биосинтез мономеров (аминокислот, нуклеотидов, моносахаров, жирных кислот) и биосинтез полимеров (белков, нуклеиновых кислот, полисахаридов и липидов). Для их синтеза необходимо около 70 различных мономеров-предшественников. Помимо них, клетка должна синтезировать ряд соединений, играющих каталитическую роль. Синтез любого мономера происходит (при наличии источников углерода и энергии) через цепь последовательных биохимических реакций, катализируемых специфическими белками-ферментами. В свою очередь синтез всех биополимеров также требует участия специфических белков. Поэтому основу основ конструктивного обмена составляет биосинтез белков, который находится под контролем генетической системы организма.