



Сергей Бабичев
Медицинская микробиология, иммунология и вирусология
Эндоспоры и спорообразование
Некоторые роды бактерий (Bacillus, Clostridium, Sporosarcina) при неблагоприятных для их существования условиях образуют защитные формы – эндоспоры. Споры представляют собой своеобразные покоящиеся клетки; у них чрезвычайно низкая метаболическая активность, но они обладают высокой устойчивостью к высушиванию, действию повышенной температуры и различных химических веществ. Высокую резистентность спор к действию указанных факторов связывают с присутствием в оболочке большого количества кальциевой соли дипиколиновой кислоты. Споры сильно преломляют свет, поэтому они хорошо заметны в неокрашенных препаратах. Для обнаружения спор предложены различные интенсивные способы окрашивания, поскольку они слабо воспринимают красители. Диаметр споры может не превышать диаметра вегетативной клетки (Bacillus) или превышает его. В последнем случае бактериальная клетка со спорой принимает форму веретена (Clostridium). Споры в клетке (рис. 15) могут располагаться центрально (B. megaterium), субтерминально (С. botulinum), терминально (C. tetani).
В процессе спорообразования (споруляции) бактериальная клетка подвергается сложной перестройке (рис. 16). Вначале на одном из ее полюсов происходит конденсация нуклеоида и отделение его за счет образования септы. Затем ЦМ начинает обрастать образовавшийся протопласт споры и возникает складка, состоящая из двух слоев ЦМ, позднее они сливаются, в результате образовавшаяся предспора оказывается окруженной двойной оболочкой. На следующей стадии между двумя мембранами, покрывающими предспору, формируется толстый слой кортекса (коры). Самый внутренний слой его представляет собой зародышевую стенку (из него образуется клеточная стенка прорастающей вегетативной клетки). По мере созревания споры обе ее мембраны участвуют в образовании специальных слоев споры. Таким образом между обращенными друг к другу мембранами образуются зародышевая стенка, кортекс, а также расположенные снаружи от мембран наружная и внутренняя оболочки и экзоспорий. Сформировавшаяся эндоспора состоит из протопласта с нуклеоидом, стенки споры, кортекса, оболочки и экзоспория.
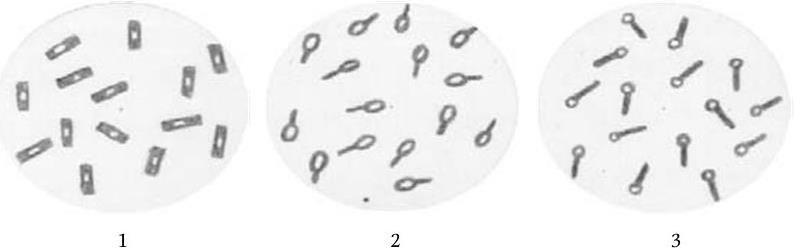
Рис. 15. Расположение спор у бактерий:
1 – центральное (Bacillus megaterium); 2 – субтерминальное (Clostridium botulinum); 3 – терминальное (Clostridium tetani)
Протопласт споры (ядро) содержит ЦМ, цитоплазму, хромосому, все компоненты белоксинтезирующей системы и анаэробной энергообразующей системы.
Стенка споры непосредственно окружает внутреннюю мембрану ее и представлена пептидогликаном, из которого формируется клеточная стенка прорастающей клетки.
Кортекс – самый толстый слой оболочки споры. Он состоит из пептидогликана, содержащего мало поперечных сшивок и поэтому очень чувствительного к лизоциму. Разрушение кортекса лизоцимом играет пусковую роль в процессе прорастания споры.
Оболочка споры построена из кератиноподобного белка. Плохая проницаемость ее определяет высокую устойчивость спор к действию различных химических веществ.
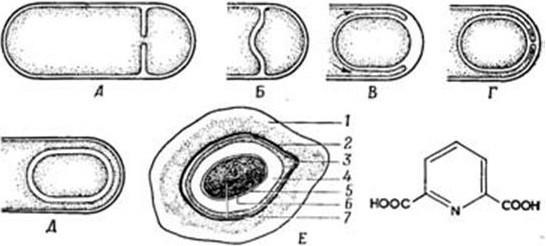
Рис. 16. Схема образования споры (по Г. Шлегелю):
А, Б – образование септы; В, Г – окружение протопласта споры мембраной материнской клетки; Д – формирование кортекса и оболочек споры; Е – схема строения зрелой споры: 1 – экзоспориум; 2 – наружная оболочка споры; 3 – внутренняя оболочка споры; 4 – кортекс; 5 – клеточная стенка споры; 6 – ЦМ споры; 7 – цитоплазма с нуклеоидом
Экзоспорий – липопротеиновая оболочка, содержащая немного углеводов.
После завершения спорообразования вегетативная часть клетки отмирает, спора высвобождается и длительное время сохраняется в окружающей среде, до тех пор, пока не возникнут условия, благоприятные для ее прорастания.
Генетический контроль спорообразования
Процесс спорообразования контролируется более чем 40 оперонами, которые представляют собой как бы дополнительный геном у спорообразующих бактерий. В составе этого генома насчитывается более 60 генов. Инициация споруляции связана с геном spoO, мутации в котором делают невозможным образование споры с самых начальных стадий. Транскрипция гена spoO запускает последовательную транскрипцию всех оперонов спорового генома. При этом их транскрипция носит строго регулируемый характер: выражение более поздних генов зависит от транскрипции более ранних генов. Это обусловливает четкую временнwую последовательность биохимических и морфологических процессов, лежащих в основе споруляции. Спорообразующие бактерии обладают механизмами, с помощью которых они распознают определенные изменения в окружающей среде, например, уменьшение содержания источников энергии, некоторых аминокислот и оснований. В ответ на это в клетке происходят метаболические изменения, которые и запускают споруляцию. Эти изменения приводят прежде всего к изменению субъединичного состава РНКполимераз. Индукция транскрипции спорового генома приводит к синтезу особых δ-единиц РНК-полимераз, которые и обеспечивают распознавание промоторов генов, контролирующих споруляцию. Вместе с тем наличие множественных промоторов у жизненно важных для клетки генов, распознаваемых разными δ-факторами, обеспечивает их выражение на всех этапах роста этих клеток, споруляции и прорастания спор. Одна из особенностей споруляции состоит в том, что на определенном ее этапе (приблизительно на 3-м часу) происходит синтез небольших кислоторастворимых белков. На их долю приходится около 10 – 12 % всех белков споры. В спорах они связываются с ДНК, обеспечивая устойчивость их к УФ-облучению. В момент прорастания споры эти белки гидролизуются и тем самым обеспечивают прорастающую спору необходимыми аминокислотами. У некоторых представителей рода Clostridium выявлена функциональная зависимость токсинообразования от споруляции: наиболее интенсивно экзотоксин вырабатывается во время активной споруляции; причинная связь этих процессов не ясна.
Прорастание споры происходит после получения соответствующего химического сигнала. Различные виды спорообразующих бактерий располагают рецепторами, распознающими наличие в среде источников энергии, L-аланина, аденозина и других веществ. Связывание с такими эффекторами активирует содержащийся в споре автолизин (лизоцим), который быстро разрушает пептидогликан кортекса.
Прорастание спор включает три стадии: активацию, начальную стадию и стадию роста.
Активация является обязательным условием прорастания спор. Она осуществляется различными воздействиями – кислой рН; веществами, содержащими свободные сульфгидрильные группы; повышением температуры; механическим повреждением спор.
Начальная стадия. Под влиянием внешних эффекторов происходит активация автолизина, последний разрушает пептидогликан кортекса, в спору поступает вода, спора высвобождается от дипиколината кальция, под воздействием гидролитических ферментов разрушаются другие ее компоненты.
Стадия роста. После разрушения кортекса и наружных слоев споры из нее появляется («выклевывается») растущая новая вегетативная клетка. Она состоит из протопласта споры и ее клеточной стенки. В ней активизируются биосинтетические процессы; в результате новая вегетативная клетка, при наличии необходимых питательных веществ, удваивает свою биомассу и делится на две дочерние клетки, которые далее активно размножаются, пока этому способствуют условия среды. Процесс прорастания споры контролируется генами как спорового, так и вегетативного геномов.
Некультивируемые формы бактерий
У многих видов грамотрицательных бактерий, в том числе у патогенных (шигеллы, сальмонеллы, холерный вибрион и др.) существует особое приспособительное, генетически регулируемое состояние, физиологически эквивалентное цистам, в которое они могут переходить под влиянием неблагоприятных условий и сохранять жизнеспособность до нескольких лет. Главная особенность этого состояния заключается в том, что такие бактерии не размножаются и поэтому не образуют колоний на плотной питательной среде. Такие не размножающиеся, но жизнеспособные клетки получили название некультивируемых форм бактерий (НФБ). Клетки НФБ, находящиеся в некультивируемом состоянии (НС), обладают активными метаболическими системами, в том числе системами переноса электронов, биосинтеза белка и нуклеиновых кислот, и сохраняют вирулентность. Их клеточная мембрана более вязкая, клетки обычно приобретают форму кокков, имеют значительно уменьшенные размеры. НФБ обладают более высокой устойчивостью во внешней среде и поэтому могут переживать в ней длительное время (например, холерный вибрион в грязном водоеме), поддерживая эндемическое состояние данного региона (водоема). Для обнаружения НФБ используют молекулярно-генетические методы (ДНК – ДНК-гибридизация, ЦПР), а также более простой метод прямого подсчета жизнеспособных клеток. С этой целью к исследуемому материалу добавляют в небольшом количестве питательные вещества (дрожжевой экстракт) и налидиксовую кислоту (для подавления синтеза ДНК) на несколько часов. Клетки усваивают питательные вещества и увеличиваются в размерах, но не делятся, поэтому такие увеличенные клетки четко видны в микроскоп и их легко подсчитать. Для этих целей можно использовать также методы цитохимические (образование формазана) или микроауторадиографии. Генетические механизмы, обусловливающие переход бактерий в НС и их реверсию из него, не ясны.
Глава 5 Физиология бактерий. Механизмы питания
Жизнь любого организма сводится к тому, чтобы в соответствии с имеющимся у него объемом генома полностью воспроизвести самого себя и реализовать свои функции, т. е. обусловить индивидуальное развитие (жизнь) и произвести потомство. Это оказывается возможным потому, что в основе жизни каждого организма лежит непрерывное взаимодействие трех потоков информации: одного – из внешней среды и еще двух потоков генетической информации: по горизонтали, он обеспечивает индивидуальное развитие организма и реализацию его жизненных функций, и другого – по вертикали, который обеспечивает передачу потомству всех признаков родителей, т. е. наследственную непрерывность вида и самой жизни. Из этого вытекает следующее положение, которое, по-видимому, имеет общебиологическую закономерность – поведение всех живых существ, как в интересах их индивидуального развития, так и ради сохранения вида, должно быть всегда адекватным имеющейся у них генетической информации и информации, воспринимаемой из внешней среды. Отступление от этого закона может привести к гибели организма и вида. Единство организма с внешней средой заключается не только в том, что он получает из среды необходимые для себя источники энергии, питания, а также другую информацию, но и в том, что, в свою очередь, он также воздействует на среду, изменяет ее и этим постоянно меняет условия своего существования. Поэтому взаимосвязь организма с внешней средой должна быть постоянной и взаимовыгодной.
Чем больше объем генома, тем сложнее устроен организм, тем разнообразнее его собственная генетическая информация и та информация, которую он может воспринимать из окружающей среды и перерабатывать. Тем разнообразнее, сложнее и богаче проявляется его индивидуальная жизнь.
Бактерии воспринимают информацию из внешней среды в виде механических, физических и, главным образом, химических сигналов, поступающих через клеточную мембрану. Химическими сигналами для бактерий служат различные источники энергии, аминокислоты, основания, другие сложные химические вещества, ионы, вода, от которых зависит общая интенсивность всех биосинтетических процессов в клетке.
Механизмы питания бактерий
Большинство бактерий живет в среде, мало подходящей для того, чтобы поддерживать строгое соотношение воды, солей и органических веществ, без которого невозможна жизнь. Это обусловливает необходимость постоянного и тщательного регулирования обмена различными веществами, который происходит между клеткой и внешней средой и контролируется клеточной мембраной. Она проницаема для многих веществ, поток их идет в обоих направлениях (из клетки и в клетку), но структура мембраны такова, что она обладает избирательной и неравномерной проницаемостью, определяющей механизмы питания бактерий.
Питательные вещества из внешней среды поступают в бактериальную клетку с помощью трех основных механизмов: пассивной диффузии, облегченной диффузии и активного транспорта (рис. 17).
Пассивная диффузия осуществляется за счет различного содержания питательных веществ в среде и в клетке и происходит в направлении от большей концентрации к меньшей, т. е. по градиенту концентрации. Когда концентрация вещества по ту и другую сторону мембраны уравнивается, пассивная диффузия прекращается. Ее скорость зависит от величины градиента, но она имеет определенный предел. Таким путем в клетку проникает (и покидает ее) вода вместе с растворенными в ней различными мелкими молекулами, способными проходить через мелкие поры мембраны. Для пассивной диффузии характерно отсутствие субстратной специфичности, и она не требует затраты энергии.
Облегченная диффузия характеризуется выраженной субстратной специфичностью и протекает при обязательном участии специфических белков, локализованных в мембране; синтез некоторых из них индуцируется соответствующими субстратами. Эти белки, получившие название пермеаз (англ. permeate – проникать, проходить сквозь), обладают субстратной специфичностью. Они распознают и связывают молекулу субстрата на внешней стороне мембраны и обеспечивают каким-то образом ее перенос через мембрану. На внутренней поверхности мембраны комплекс пермеаза – субстрат диссоциирует, освободившаяся молекула субстрата включается в общий метаболизм клетки, а пермеаза повторяет очередной цикл переноса своего субстрата, который не способен проникать через мембрану путем простой диффузии. Главное свойство пермеаз – способность проходить через мембрану как с присоединенной молекулой субстрата, так и без нее. Однако облегченная диффузия происходит только по градиенту концентрации, но не против него, поэтому она не требует затраты энергии. Пермеазы, присоединившись к субстрату, повышают его способность проникать через мембрану. Облегченная диффузия протекает со значительно более высокой скоростью, чем пассивная. Ее скорость подчиняется закону Михаэлиса – Ментен, и при достижении равновесия концентрация субстрата, доставляемого посредством облегченной диффузии, на внутренней и внешней поверхностях мембраны становится одинаковой.
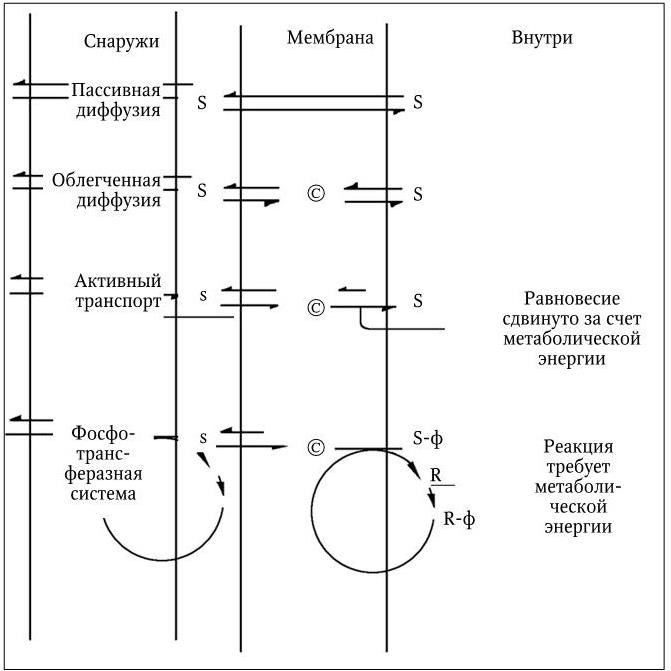
Рис. 17. Бактериальные транспортные системы (Р. Стейнер [и др.]. Мир микробов. 1979, т. 2):
Разная длина стрелок указывает на сдвиг равновесия реакции в сторону более длинной стрелки. S и s означают соответственно высокую и низкую концентрации растворенных веществ; © – белок-переносчик (пермеаза); R – белок HРr; R-ф – фосфо-HРr; ф – фосфатная группа
Активный транспорт. С помощью механизмов активного транспорта растворенные вещества могут поступать в клетку против градиента концентрации, поэтому активный транспорт требует от клетки затраты энергии. У бактерий этот механизм питания является преобладающим. С его помощью они обеспечивают такие концентрации растворенных питательных веществ внутри клетки, которые могут во много раз превышать их концентрации во внешней среде и обеспечивают им высокие скорости метаболизма даже при низкой концентрации химических веществ в окружающей среде. У многих бактерий, в особенности грамотрицательных, в активном транспорте принимают участие особые связывающие белки, не идентичные пермеазам и не входящие в структуру мембраны, а локализованные в периплазматическом пространстве. У связывающих белков отсутствует каталитическая активность, но они обладают очень высоким сродством к определенным питательным веществам – к различным аминокислотам, сахарам, неорганическим ионам. Выделено и изучено более 100 различных связывающих белков, которые образуют прочные комплексы со своими субстратами и необходимы для их активного переноса через мембрану. Связывающие белки функционируют только в комплексе со специфическими пермеазами, осуществляющими активный перенос субстрата через мембрану. Метаболическая энергия, необходимая для этого, используется для снижения сродства пермеазы к своему субстрату на внутренней поверхности мембраны по сравнению с ее сродством к нему на внешней стороне мембраны. В результате этих превращений происходит изменение скорости выхода субстрата наружу, она становится во много раз меньше скорости его поступления в клетку. При этом механизме активного транспорта через мембрану в клетку поступают против градиента концентрации химически не измененные питательные вещества. У бактерий, вместе с тем, существуют и такие транспортные системы, которые переводят питательные вещества в химически измененную форму, не способную проникать через мембрану. К их числу относится фосфотрансферазная система, широко распространенная среди бактерий. С помощью этой системы транспортируются многие сахара и их производные, в процессе переноса они фосфорилируются и поступают в клетку в виде сахарофосфатов. Поскольку мембрана для последних непроницаема, сахарофосфаты остаются внутри клетки.
Фосфотрансферазная система состоит из двух неспецифических компонентов: ферментов I и HPr и набора субстрат-специфических белков, связанных с мембраной и обозначенных как ферменты II. Фермент I обеспечивает перенос богатой энергией фосфатной группы от фосфоенолпирувата на гистидиновый остаток фермента HPr, который превращается в фосфо-HPr. Последний является общим донором фосфорильной группы для всех субстратов, переносимых фосфотрансферазной системой. Фосфорилирование же их осуществляется субстрат-специфическими белками из группы ферментов II, которые выполняют также и функции пермеаз. У мутантных бактерий, лишенных фермента I или белка HPr, ферменты II осуществляют облегченную диффузию своих субстратов.
Транспортные системы в жизни клетки выполняют две основные функции:
1) поддерживают на высоком уровне внутриклеточные концентрации всех субстратов, необходимых для осуществления важнейших биохимических реакций с максимальными скоростями даже при низких концентрациях этих химических веществ во внешней среде;
2) регулируют внутриклеточное осмотическое давление, поддерживают оптимальную для метаболической активности концентрацию растворенных веществ (небольших молекул и ионов).
Секреция продуктов жизнедеятельности бактериальной клеткой
Бактерии синтезируют и секретируют во внешнюю среду различные продукты своей жизнедеятельности, в том числе белки, главным образом ферменты и токсины, с помощью которых они оптимизируют свое существование. Например, благодаря секреции ферментов они расщепляют сложные органические соединения и делают их более доступными для питания. Для патогенных бактерий секреция ферментов и токсинов служит мощным средством, благодаря которому они только и могут обеспечивать свое существование и размножение в организме животного и подавлять его защитные механизмы. Секреция белков бактериями осуществляется с помощью различных систем и механизмов. При этом следует различать секрецию белков в периплазматическое пространство через ЦМ и секрецию белков непосредственно в культуральную среду (экскрецию, или экспорт белков). У грамотрицательных бактерий большинство белков секретируется в периплазматическое пространство в виде белков-предшественников, содержащих в своей структуре особый сигнальный (лидерный) пептид из 15 – 40 аминокислотных остатков. Именно он обеспечивает перенос белка-предшественника через ЦМ, после чего и отрезается от него с помощью сигнальной (лидерной) пептидазы.
Существует несколько моделей, объясняющих механизм, посредством которого лидерный пептид обеспечивает секрецию белка-предшественника через ЦМ в периплазматическое пространство.
Модель прямого транспорта предполагает прямое вхождение лидерного пептида в липидный бислой мембраны с использованием свободной энергии мембраноассоциированных рибосом.
Сигнальная гипотеза предполагает, что в результате взаимодействия лидерного пептида непосредственно с особым рецептором мембраны образуется внутримембранный канал, через который и осуществляется секреция.
Существуют и другие, более сложные, модели механизма переноса секретируемого белка через ЦМ. Возможно, что применительно к разным белкам и у разных групп бактерий действуют различные механизмы. Детальнее всего механизмы секреции изучены у E. coli. У нее обнаружены два пути секреции: sec-зависимый и относительно sec-независимый. Для обеспечения секреции белков в случае sec-зависимого механизма требуется участие продуктов ряда sec-генов: secA, secY, secB, secD, secE и secF. Источниками энергии для переноса белков служат гидролиз АТФ и градиент концентрации протонов. Для осуществления посттранслокационного процессинга (англ. processing – обработка) белка после его перемещения (транслокации) достаточно, вероятно, активности только сигнальных пептидаз. У E. coli их обнаружено две: сигнальная пептидаза I (м. м. 36 кД, кодируется геном lepB) и сигнальная пептидаза II (м. м. 18 кД, кодируется геном lepA).
Большой интерес представляет так называемая система секреции 3-го типа (ССТ3). Она осуществляет секрецию эффекторных белков из цитоплазмы клетки через ЦМ и наружную мембрану непосредственно в клетки растения и животного, с которыми бактерия контактирует. ССТ3 обнаружена у бактерий родов Shigella, Salmonella, Yersinia и других и играет у них роль одного из факторов патогенности. Непосредственно в культуральную среду грамотрицательные бактерии экскретируют только некоторые белки, при этом в каждом случае используются различные механизмы, которые также еще недостаточно изучены. Например, бактериоцины, кодируемые различными плазмидами, не содержат в своей структуре сигнальных пептидов. Для их секреции через ЦМ и наружную мембрану требуется специальный вспомогательный белок – рилизинг-белок. Система транспорта гемолизина HlyA, кодируемого генами Hly-плазмиды, состоит как минимум из двух вспомогательных белков HlyB и HlyD, которые образуют канал для непосредственного выхода гемолизина (важного фактора патогенности E. coli) из цитоплазмы во внешнюю среду.