



Уильям Бейтс
Совершенное зрение без очков. Лечение несовершенного зрения без помощи очков
Глава IV. Правда об аккомодации. Демонстрация во время экспериментов с глазными мышцами рыб, кошек, собак, кроликов и других животных
Функция мышц, находящихся на внешней части глазного яблока, помимо вращения глаза в глазнице, стала причиной большого количества споров. Но после мнимой демонстрации Гельмгольцем того, что аккомодация зависит от изменения кривизны хрусталика, их возможное предназначение настраивать глаз на работу на различные расстояния или их участие в создании аномалий рефракции было отвергнуто и больше не считалось достойным какого-либо внимания. «Прежде чем физиологи ознакомились с изменениями в диоптической системе[45], – говорит Дондерс, – они часто приковывали свое внимание к внешним мышцам глаза в процессе совершения аккомодации. Сейчас, когда мы знаем, что аккомодация зависит от формы хрусталика, нет оснований опровергать данную точку зрения». Он решительно заявляет о том, что «наблюдается много случаев, когда аккомодация полностью парализована без какого-либо воспрепятствования этому со стороны внешних мышц», а также что «во многих зарегистрированных случаях паралича всех или практически всех мышц глаза, а также при отсутствии этих мышц ослабления способности к аккомодации не наблюдалось».[46]
Если бы Дондерс не счел этот вопрос решенным, он, возможно, изучил бы более скрупулезно все эти случаи. И если бы это произошло, то он мог бы быть менее категоричен в своих заявлениях, потому что, как было показано в предыдущей главе, существует огромное множество признаков, указывающих на то, что на самом деле все происходит с точностью до наоборот. В моих собственных экспериментах с внешними мышцами глаз рыб, кроликов, кошек, собак и других животных было полностью продемонстрировано, что в глазах этих животных аккомодация целиком зависит от деятельности внешних мышц и происходит безо всякого участия хрусталика. Производя манипуляции с этими мышцами, я смог по своему усмотрению воспроизводить или не давать происходить аккомодации, воспроизводить миопию, гиперметропию и астигматизм или же предотвращать возникновение этих состояний. Полное описание этих экспериментов вы можете найти в «Бюллетене Нью-Йоркского зоологического общества» за ноябрь 1914 года и в «Нью-Йоркском медицинском журнале» за 8 мая 1915 года и за 18 мая 1918 года. Но для тех, кто не имеет времени или возможности прочитать эти издания, их содержание я описал ниже.
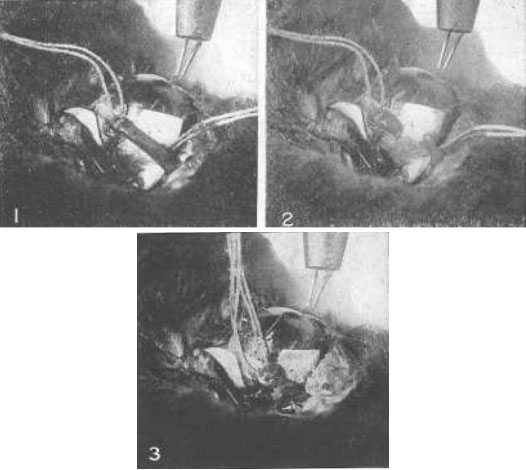
Рис. 13. Демонстрация на примере глаза кролика того, что нижняя косая мышца является существенным фактором в процессе аккомодации
№ 1 – Нижняя косая мышца была подвержена воздействию электрическим током, и две нити были присоединены к ней. Стимуляция глазного яблока электрическим током вызвала аккомодацию, как показала симультативная ретиноскопия.
№ 2 – Мышца была разрезана. Стимуляция электрическом током не вызывает аккомодации.
№ 3 – Концы разрезанной мышцы сшиты друг с другом. Стимуляция электрическим током способствует возникновению нормальной аккомодации
Существует шесть мышц, находящихся на внешней части глазного яблока, четыре из которых известны как «прямые», а две другие – как «косые». Косые мышцы практически полностью опоясывают глазное яблоко посередине, и в соответствии с их расположением они также известны как «верхние» и «нижние». Прямые мышцы присоединены к склеротической, или внешней, оболочке глазного яблока, ближе к передней его части, и идут, минуя верх, низ и боковые части глазного яблока, прямо до задней части глазницы, где они присоединяются к костным тканям по краям круглого отверстия, через которое проходит зрительный нерв. В соответствии с их расположением они носят названия «верхних», «нижних», «внутренних» и «внешних» прямых мышц. Косые мышцы – это мышцы аккомодации, прямые воспроизводят гиперметропию и астигматизм.
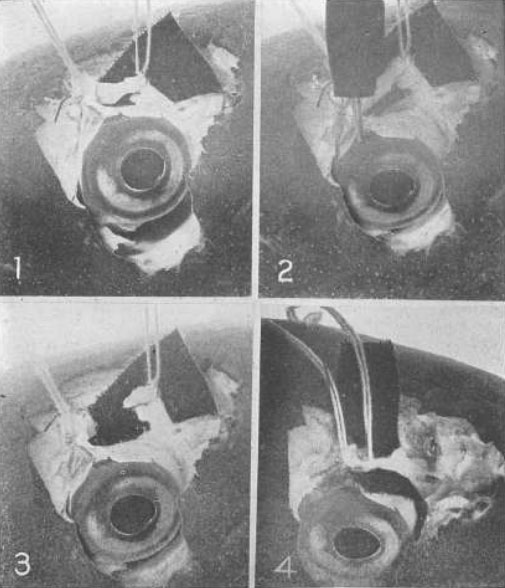
Рис. 14. Демонстрация на примере глаза карпа того, что верхняя косая мышца играет значимую роль в процессе аккомодации
№ 1 – Верхняя косая мышца оттянута вверх от глазного яблока при помощи двух нитей, и ретиноскоп показывает отсутствие аномалий рефракции. № 2 – Как определил ретиноскоп, стимуляция электрическим током приводит к возникновению аккомодации. № 3 – Мышца была разрезана. Стимуляцией глазного яблока электрическим током не удалось воспроизвести аккомодацию. № 4 – Разрезанная мышца вновь была соединена путем сшивания. Как и прежде, в результате стимуляции электрическим током возникает аккомодация.
В некоторых случаях одна из косых мышц отсутствует или неразвита. Но когда две эти мышцы присутствовали и могли действовать, аккомодация, как было измерено при помощи объективного теста в виде ретиноскопии, всегда происходила под действием стимуляции электрическим током либо глазного яблока, либо нервов аккомодации возле участка в головном мозге, откуда они выходили. Аккомодация также воспроизводилась путем любой манипуляции с косыми мышцами, в результате которой усиливалось их натяжение. Такое натяжение осуществлялось при помощи операции по подворачиванию одной или обеих мышц (англ. «tucking operation». – Прим. пер.) или путем увеличения расстояния до точки, в которой они присоединялись к склере. Когда одна или несколько прямых мышц были разрезаны, эффект от операции, заключавшийся в увеличении натяжения косых мышц, был усилен.
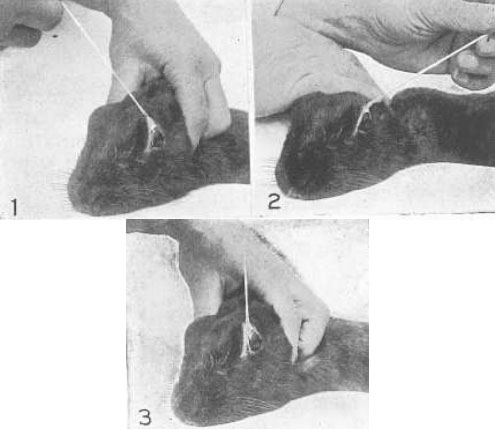
Рис. 15. Демонстрация на примере глаза кролика того, что возникновение аномалий рефракции зависит от деятельности внешних мышц глаза. Шнур привязан к месту крепления верхней косой и прямой мышц глаза
№ 1 – Нить тянется назад. Возникает миопия.
№ 2 – Нить тянется вперед. Возникает гиперметропия.
№ 3 – Нить тянется вверх в плоскости радужки. Возникает смешанный астигматизм.
После того как были разрезаны поперек одна или обе косые мышцы или после того как их парализовывало в результате инъекции атропина глубоко в глазницу, аккомодация никогда не вызывалась при помощи стимуляции электрическим током. Но после окончания действия атропина или когда разделенные концы мышцы сшивались друг с другом, за электрическим разрядом, как и обычно, следовала аккомодация. И вновь, когда одна косая мышца отсутствовала, как было обнаружено в случае морской собаки, акулы и нескольких окуней, или была неразвита, как в случаях всех исследованных кошек, нескольких рыб и кролика, не удавалось воспроизвести аккомодацию при помощи стимуляции электрическим током. Но когда неразвитая мышца была усилена дополнительным удлинением или отсутствовавшую мышцу заменял шнур, поддерживавший необходимое натяжение, всегда удавалось воспроизвести аккомодацию при помощи электрического тока.

Рис. 16. Демонстрация на глазу рыбы того, что воспроизведение миопической и гиперметропической рефракций зависит от действия внешних мышц
Шнур привязан к основанию верхней прямой мышцы. Сильно потянув за конец шнура, повернули глазное яблоко в глазнице, и, затягивая нить при помощи фиксирующего зажима, захватывающего нижнюю челюсть, его устанавливают в этом положении. Симультативная ретиноскопия зарегистрировала воспроизведение высокой степени смешанного астигматизма. Когда верхняя косая мышца разделена, миопическая составляющая астигматизма исчезает, а когда разрезана нижняя прямая мышца, то гиперметропическая составляющая исчезает и глаз становится нормальным – настроенным на зрение вдаль, – хотя поддерживается натяжение шнура той же силы. Тем доказано, что эти мышцы являются существенными факторами в создании миопии и гиперметропии.
После того как одна или обе косые мышцы были разрезаны и в то время как две и более прямых мышц присутствовали и были активны[47], стимуляция глазного яблока или нервов аккомодации электрическим током всегда воспроизводила гиперметропию. В то же время при манипуляциях с одной из прямых мышц – обычно нижней или верхней, – так чтобы усилить их натяжение, получался такой же результат. Парализация прямой мышцы при помощи атропина или разрезание одной или нескольких таких мышц не позволяли возникнуть гиперметропической рефракции при электрической стимуляции. Но после окончания действия атропина или после того, как разрезанные концы мышцы были сшиты друг с другом, как обычно, в результате стимуляции электрическим током возникала гиперметропия.
Следует подчеркнуть, что, для того чтобы парализовать либо прямые, либо косые мышцы, оказалось необходимым производить инъекции атропина далеко позади глазного яблока при помощи иглы для подкожных инъекций. Предполагалось, что препарат парализует аккомодацию, когда его закапывают в глаза людей или животных, но во всех моих экспериментах было обнаружено, что, когда его использовали таким образом, он оказывал очень небольшой эффект на способность глаза изменять свой фокус.
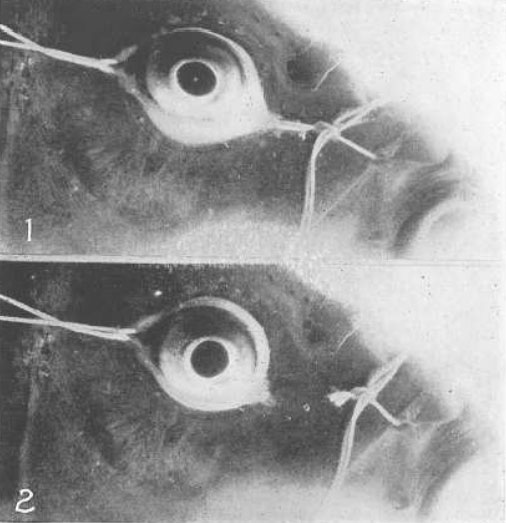
Рис. 17.
№ 1 – Воспроизведение смешанного астигматизма в глазу карпа путем оттягивания нитей, прикрепленных к конъюнктиве, в противоположных направлениях. Заметьте овальную форму передней части глазного яблока.
№ 2 – После перерезания нитей глазное яблоко возвращает свою нормальную форму и рефракция становится нормальной.
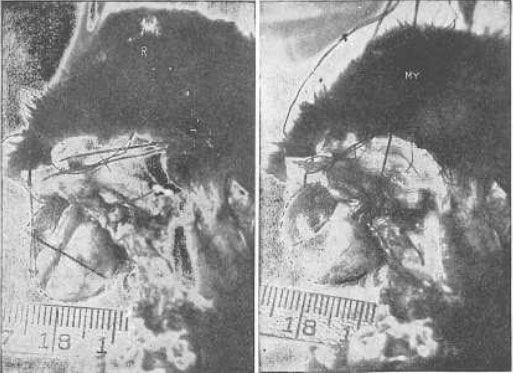
Рис. 18. Демонстрация на глазном яблоке кролика того, что косые мышцы удлиняют зрительную ось при миопии
R, состояние покоя. Глазное яблоко имеет нормальную длину и находится в состоянии эмметропии – то есть полностью настроено на зрение вдаль. Му, миопия. Натяжение косых мышц было усилено путем их перемещения, и ретиноскоп показал, что возникла миопия. Легко заметить, что глазное яблоко стало длиннее. Было невозможно избежать какого-либо движения головы между съемкой этих двух кадров, на которых изображены результаты манипуляции с нитями. Но линейка показывает, что фокус камеры не был сильно изменен такими перемещениями.
Астигматизм обычно возникал в комбинации с миопической или гиперметропической рефракциями. Его также удавалось воспроизводить при помощи различных манипуляций и с косыми, и с прямыми мышцами. Смешанный астигматизм, который является комбинацией миопической и гиперметропической рефракций, всегда воспроизводился при натяжении в местах крепления верхней или нижней прямых мышц в направлении, параллельном плоскости радужки, при условии, что обе косые мышцы присутствовали и могли действовать. Но если одна или обе косые мышцы были разрезаны, миопическая составляющая астигматизма исчезала. Подобным образом после разрезания верхней или нижней прямых мышц исчезала гиперметропическая составляющая астигматизма. Перемещение двух косых мышц с перемещением верхней и нижней прямых мышц всегда воспроизводило смешанный астигматизм.
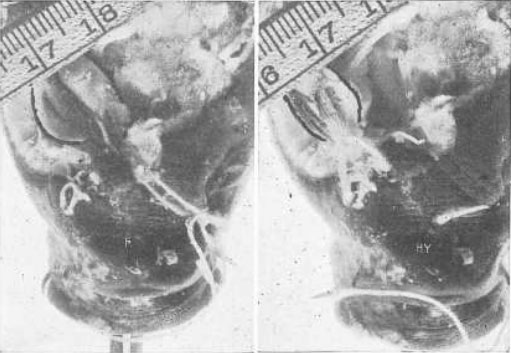
Рис. 19. Демонстрация на примере глаза карпа того, что прямые мышцы укорачивают зрительную ось при гиперметропии
R, состояние покоя. Глазное яблоко имеет нормальную длину и находится в состоянии эмметропии. Ну, гиперметропия. Натяжение внешней и внутренней прямых мышц было усилено путем перемещения, и ретиноскоп показывает, что возникла гиперметропия. Еще легче заметить, что глазное яблоко стало короче. Линейка показывает, что фокус камеры не был существенно изменен между этими двумя фотографиями.
Глаза, из которых был удален хрусталик или в которых он был смещен со зрительной оси, реагировали на стимуляцию электрическим током точно так же, как это делали нормальные глаза, все то время, что мышцы были активны. Но когда они были парализованы инъекцией атропина глубоко в глазницу, стимуляция не оказывала никакого влияния на рефракцию.

Рис. 20. Хрусталик смещен со зрительной оси.
В этом эксперименте на глазу карпа хрусталик был вытеснен со зрительной оси. Аккомодация имеет место после этого смещения точно так же, как и в предыдущих случаях. Заметьте точку на ноже в зрачке на передней поверхности хрусталика.
В одном из экспериментов из правого глаза кролика был удален хрусталик. Сначала рефракция каждого глаза была проверена при помощи ретиноскопа и оказалась нормальной. Потом ранке дали время зажить. После этого в период времени, начиная от одного месяца и до двух лет, в глазу с удаленным хрусталиком всегда удавалось воспроизвести аккомодацию при помощи стимуляции электрическим током в том же объеме, что и в глазу, имевшем хрусталик. Тот же эксперимент с таким же результатом был проведен на нескольких других кроликах, на собаках и на рыбах. Очевидный вывод – хрусталик не является фактором в процессе аккомодации.
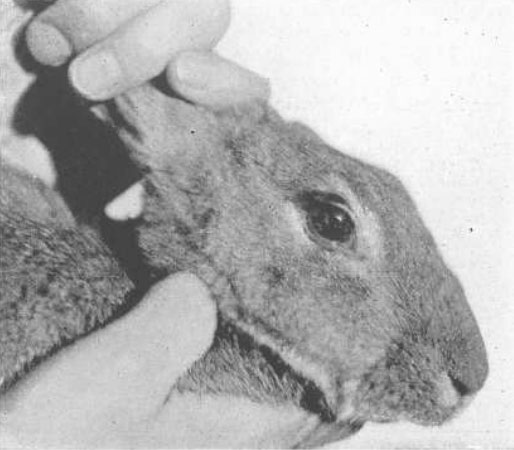
Рис. 21. Кролик с удаленным хрусталиком
Это животное было представлено на заседании офтальмологов Американской ассоциации, проведенном в Атлантик-Сити, и было исследовано несколькими присутствовавшими там офтальмологами, все из которых свидетельствовали о том, что стимуляция глазного яблока электрическим током приводит к возникновению аккомодации или миопической рефракции точно так же, как и в нормальном глазу.
В большинстве учебников по физиологии написано, что аккомодация контролируется третьим черепным нервом, который снабжает все мышцы глазного яблока за исключением верхней косой и внешней прямой мышц. Но в этих экспериментах было обнаружено, что четвертый черепной нерв, который снабжает только верхнюю косую мышцу, – такой же нерв аккомодации, как и третий. Когда либо третий, либо четвертый нерв был стимулирован электрическим током возле точки его выхода из головного мозга, в нормальном глазу всегда возникала аккомодация. Когда начало каждого нерва было покрыто маленьким кусочком ваты, смоченным двухпроцентным раствором атропина сульфата в нормальном физиологическом растворе, стимуляция этого нерва не приводила к возникновению аккомодации, тогда как стимуляция не парализованного нерва воспроизводила ее. Когда основания обоих нервов были покрыты ватой, смоченной в атропине, аккомодацию не удавалось получить путем стимуляции электрическим током одного из них или обоих нервов. Когда вата была удалена и нервы промыты нормальным физиологическим раствором, стимуляция одного или обоих нервов электрическим током воспроизводила аккомодацию точно так же, как и до применения атропина. Этот эксперимент, проведенный повторно в течение более одного часа попеременным применением и удалением атропина, не только ясно продемонстрировал то, о чем ранее не было известно, а именно то, что четвертый нерв является нервом аккомодации, но также продемонстрировал, что верхняя косая мышца, которая его снабжает, является значимым фактором в процессе аккомодации. Далее было обнаружено, что когда не давали действовать косым мышцам путем их рассечения, стимуляция третьего нерва воспроизводила не аккомодацию, а гиперметропию.
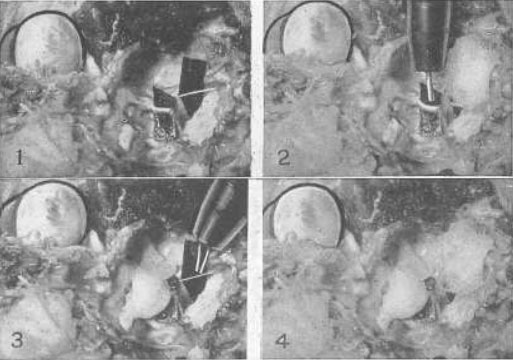
Рис. 22. Эксперимент на примере глаза кошки, демонстрирующий то, что четвертый нерв, который снабжает только верхнюю косую мышцу, является точно таким же нервом аккомодации, как и третий, и что верхняя косая мышца, которую он снабжает, есть мышца аккомодации
№ 1 – Оба нерва были выведены наружу возле их основания в головном мозге, и полоска черной бумаги была вложена позади каждого, для того чтобы изображение можно было увидеть. Четвертый нерв – это тот, что поменьше. Верхняя косая мышца была перемещена путем подгибания. Так как эта мышца всегда недоразвита у кошек, то до тех пор, пока ее натяжение не усиливают, аккомодация у этих животных не воспроизводится. Стимуляция одного или обоих нервов импульсным током способствовала возникновению аккомодации.
№ 2 – Когда четвертый нерв был покрыт ватой, пропитанной нормальным физиологическим раствором, применение импульсного тока вызвало аккомодацию. Когда вата была пропитана однопроцентным раствором атропина сульфата в нормальном физиологическом растворе, не удавалось воспроизвести аккомодацию, но стимуляция третьего нерва воспроизводила ее.
№ 3 – Когда третий нерв был покрыт ватой, пропитанной нормальным физиологическим раствором, воздействием импульсного тока была воспроизведена аккомодация. Когда вата была смочена атропина сульфатом в нормальном физиологическом растворе, то не удавалось воспроизвести аккомодацию, но стимуляция четвертого нерва аккомодацию все же вызывала.
№ 4 – Когда оба нерва были покрыты ватой, пропитанной атропина сульфатом в нормальном физиологическом растворе, применение электричества к вате не вызывало возникновения аккомодации. Когда органы были вымыты в теплом физиологическом растворе, стимуляция электрическим током каждого нерва всегда воспроизводила аккомодацию. Нервы попеременно покрывались ватой, смоченной атропином, а затем промывались теплым физиологическим раствором за один час до применения электрического тока в каждом из состояний с теми же результатами. Аккомодация никогда не могла быть воспроизведена при помощи стимуляции электрическим током, когда нервы были парализованы атропином, но всегда происходила при стимуляции каждого или обоих нервов, когда их промывали физиологическим раствором. Эксперимент был проведен с теми же результатами, что и с множеством кроликов и собаками.
Можно с уверенностью сказать о том, что во всех экспериментах все источники ошибок были исключены. Эксперименты были проведены повторно много раз и всегда – с тем же результатом. Поэтому в их правдивости нет причин сомневаться, а именно в том, что ни хрусталик, ни какая-либо мышца внутри глазного яблока не имеют ничего общего с аккомодацией, а процесс настройки глаза на зрение на различные расстояния полностью контролируется действием мышц, находящихся на внешней части глазного яблока.
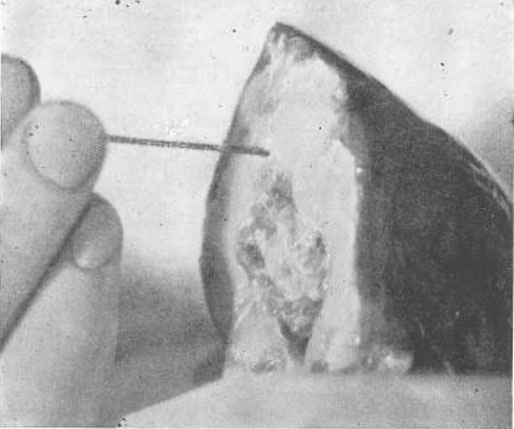
Рис. 23. Разрушение спинного мозга рыбы для подготовки к операции на ее глазах
Эта операция производится для того, чтобы установить максимальную релаксацию мышц глаз и головы, которые будут работать в течение нескольких часов без воздействия извне, если клетки головного мозга не разрушены зондом.
Глава V. Правда об аккомодации. Демонстрация в процессе изучения изображений отраженных от хрусталика, роговицы, радужки и склеры
Выводы, полученные из опытов, описанных в предыдущей главе, были диаметрально противоположны тем выводам, которые получил Гельмгольц при изучении изображений, отраженных от передней поверхности хрусталика. Поэтому я решил повторить эксперименты немецкого исследователя и по возможности найти объяснение, почему его результаты так сильно отличались от моих. Я посвятил этой работе четыре года и смог продемонстрировать, что Гельмгольц ошибся, выбрав неправильную технику проведения эксперимента: изображение, полученное его методом, было настолько изменяющимся и неясным, что с его помощью можно было подтвердить практически любую возможную теорию. Я работал год или дольше с техникой Гельмгольца, но мне так и не удалось получить изображения с передней поверхности хрусталика, которое было бы достаточно ясным или достаточно разборчивым для того, чтобы его можно было измерить или сфотографировать. Используя огонь свечи в качестве источника света, можно было получить чистое и ясное изображение с роговицы; на задней поверхности хрусталика оно было достаточно ясным; но на передней поверхности оно было очень далеким от совершенства. Как констатировал сам Гельмгольц, оно было не только размытым, но и безо всякой на то причины сильно изменялось в размере и интенсивности. Порой совсем никакого изображения не удавалось получить вне зависимости от того, под какими углами ни располагался бы свет по отношению к глазу исследуемого или к глазу исследователя. Используя диафрагму, я смог получить более ясное и более устойчивое изображение, но оно по-прежнему не поддавалось замеру. Гельмгольцу казалось, что нечеткие изображения открытого пламени свечи указывали на существенное изменение, тогда как изображения, полученные с помощью диафрагмы, показывали это более ясно; но мне так и не удалось ни с помощью диафрагмы, ни без нее получить какого-либо изображения приемлемой для меня четкости.

Рис. 24. Установка для фотографирования изображений, отраженных от глазного яблока
СМ, вогнутое зеркало, в которое исследуемая может наблюдать изображения, отраженные от различных участков ее глаз; С, конденсор; D, диафрагма; L, 1000-ваттная лампа; F, упор для лба; МР, перекладина, за которую исследуемая держится зубами, для того чтобы ее голова оставалась неподвижной; Р, плоское зеркало, над которым расположена буква шрифта «диамант» и в котором отражается таблица Снеллена, находящаяся позади исследуемой на расстоянии двадцати футов (зеркало – чуть выше Р); САМ, фотоаппарат; Pr, периметр, используемый для измерения угла между светом и глазом; R, плоское зеркало, отражающее свет от 1000-ваттной лампы над глазом: без этого зеркала глаз будет находиться в полной темноте, за исключением той его части, от которой будет отражаться сильно сжатое изображение нити накала; В, экран из голубого стекла, используемый для легкого приглушения света, отраженного от зеркала R. Когда исследуемая читала нижнюю строку таблицы Снеллена, отраженную в зеркале Р, ее глаз находился в покое, но когда она увидела букву шрифта «диамант» четко, ретиноскоп зарегистрировал аккомодацию в десять диоптрий.
Люди, преподававшие и демонстрировавшие теорию Гельмгольца, повторили для меня его эксперименты; но изображения с передней поверхности хрусталика, полученные ими, не показались мне лучше тех, что получил я сам. После тщательно изучения этих изображений почти ежедневно в течение более чем одного года я так и не смог сделать какого-либо приемлемого заключения на предмет эффекта аккомодации. В действительности казалось, что, используя свечу в качестве источника освещения, по изображению на передней поверхности хрусталика можно было наблюдать проявления бесчисленного множества различных явлений. Иногда во время аккомодации изображение уменьшалось в размере и, казалось, удовлетворяло теории Гельмгольца; но, с другой стороны, столь же часто оно увеличивалось. В другие разы было просто невозможно как-либо интерпретировать то, что происходило с этим изображением.
Применение 30-ваттной, 50-ваттной, 250-ваттной и 1000-ваттной ламп не помогло получить более качественных изображений. Солнечный свет, отраженный от передней поверхности хрусталика, создавал такое же неясное изображение, как и отражения от других источников освещения, и оно было такой же изменяющейся формы, интенсивности и размера. В результате всего описанного выше я убедился в том, что передняя поверхность хрусталика – очень плохой отражатель света и ни один из вышеуказанных способов не позволяет получить на ней приемлемых изображений.

Рис. 25. Установка для обеспечения головы исследуемого объекта в неподвижном состоянии во время фотографирования изображения
СМ, вогнутое зеркало; F, упор для лба; С, конденсор; МР, перекладина для обеспечения неподвижности головы исследуемого объекта; Pr, периметр.
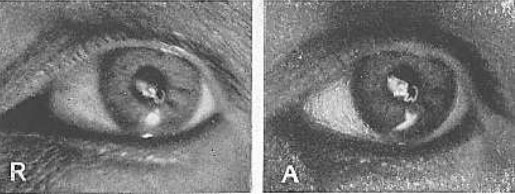
Рис. 26. Изображение электрической нити накала на передней поверхности хрусталика
R, состояние покоя; А, аккомодация. Под увеличительным стеклом никакого изменения размера двух изображений не наблюдается. Изображение справа выглядит больше только потому, что оно более четкое. Для поддержания теории Гельмгольца оно должно быть меньше. Хвостик кометы на рисунке слева – это случайное отражение от роговицы. Отблеск света внизу – просто отражение вспышки фотокамеры, на которую был запечатлен глаз. Потребовалось два года, для того чтобы получить эти фотографии.
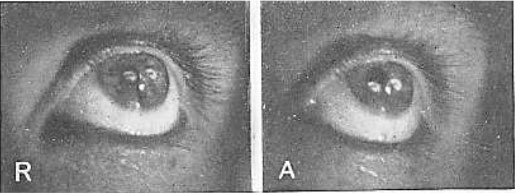
Рис. 27. Изображения электрической нити накала, одновременно отраженной от роговицы и хрусталика
R, состояние покоя; А, аккомодация. Размеры изображений на обеих картинках одинаковые. Изображение на роговице настолько мало́, что оно не претерпело заметных изменений при несильном изменении, которое имело место в роговице во время аккомодации. На рисунке А оба изображения изменили свое расположение, а замыкающая часть отражения от хрусталика была отрезана радужкой, но она остается той же самой. Белый блик между двумя изображениями нити накала есть отражение от лампы, использовавшейся для освещения глаза. Заметьте, что на рисунке А видна бо́льшая часть склеры, что указывает на удлинение глазного яблока во время аккомодации.
После более года неудачных экспериментов я начал работать в аквариуме, исследуя глаза рыб. Очень долго у меня ничего не получалось. Но наконец мне удалось при помощи очень яркого света – 1000 ватт, – диафрагмы с маленьким отверстием и конденсора получить, хоть и не без труда, но ясное и четкое изображение с роговицы рыбы. Это изображение было достаточно четким для того, чтобы его можно было измерить, и спустя несколько месяцев мне удалось получить приемлемую фотографию. Затем я снова продолжил работать с глазом человека. Яркий свет в сочетании с диафрагмой и конденсором, использование которых было предложено для улучшения освещения предметного стекла под микроскопом, как и предполагалось, оказался гораздо лучше той методики, которую использовал Гельмгольц, и с помощью этих средств было получено изображение с передней поверхности хрусталика, которое было достаточно ясным и достаточно четким для того, чтобы его можно было сфотографировать. Согласно опубликованным ранее записям, изображение какого-либо вида впервые было сфотографировано с передней поверхности хрусталика. Профессиональные фотографы, с которыми я консультировался и надеялся на их помощь, уверили меня в том, что это сделать невозможно, и отказались даже попробовать это сделать. Поэтому мне пришлось самому учиться фотографировать – с нуля, потому что у меня совсем не было опыта фотографирования. После этого мне стало ясно, что профессионалы были правы в том, что невозможно получить снимок, пользуясь методикой Гельмгольца.
Я продолжил эксперименты и проводил их до тех пор, пока спустя почти четыре года постоянных поисков не получил удовлетворительных снимков до и после аккомодации и в процессе воспроизведения миопии и гиперметропии. Не только изображений на передней поверхности хрусталика, но и отражений от радужки, роговицы, передней поверхности склеры (белая часть глаза) и от боковой поверхности склеры. У меня также стало удаваться получать изображения на любой поверхности без отражения от других частей. Однако на моем пути было еще много трудностей, которые было необходимо преодолеть.
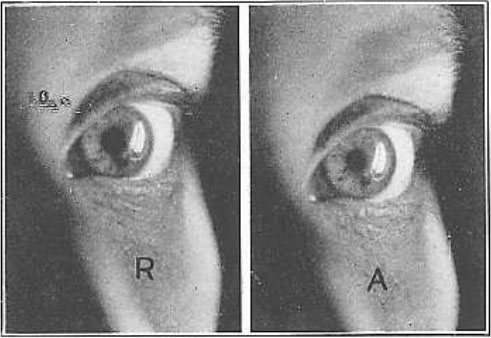
Рис. 28. Изображение электрической нити накала на поверхности роговицы
R, состояние покоя; А, аккомодация. Изображение на рисунке А меньше, но изменение настолько несущественное, что заметить его чрезвычайно трудно. Это указывает на то, что роговица лишь незначительно изменяет свою форму во время аккомодации. По этой причине думают, что офтальмометр с его маленькими изображениями демонстрирует, что форма роговицы не изменяется при аккомодации.
Накладывающиеся друг на друга отражения были нескончаемым источником проблем. Появление отражений от окружающих объектов можно было легко предотвратить; но трудно было работать с отражениями электрического света от глазного яблока, и было просто бесполезно пытаться получить изображения на передней поверхности хрусталика до тех пор, пока не удавалось устранить эти отражения или минимизировать их как только это было возможно с помощью надлежащей регулировки света. Однако регулировка освещения, которая казалась наиболее приемлемой, не всегда давала одинаковые результаты. Иногда случалось так, что не появлялось вообще никаких отражений в течение нескольких дней; но наступал день, когда свет, как я предполагаю, падал под тем же углом и изображения появлялись вновь.
С определенными настройками света можно было увидеть изображения, многократно отраженные от передней поверхности хрусталика. Иногда эти изображения были выстроены в горизонтальную линию, иногда – в вертикальную, а иногда – под различными углами, тогда как расстояния между ними также изменялись. Обычно было три изображения, иногда их было больше, а когда-то появлялось только два. Иногда все они были одного размера, но, как правило, изменялись, и казалось, что не было предела их способности изменяться в этом и в других отношениях. Какие-то из них были сфотографированы, что говорит о том, что это были реальные отражения. Изменение расстояния между диафрагмой, источником света и конденсором, а также изменение размера и формы отверстия не давали результатов, отличных от прежних. Различные настройки конденсора также не оказывали никакого эффекта. Изменение угла, под которым настраивался свет, иногда приводило к уменьшению количества изображений, но иногда их число увеличивалось. Это продолжалось до тех пор, пока наконец угол, под которым не было видно ничего, кроме одного изображения, не был найден. В действительности оказалось, что изображения были обусловлены отражениями электрического света от глазного яблока.
Даже после того как свет был настроен так, чтобы не возникало отражений, все равно было сложно, даже невозможно, получить ясное, четкое изображение электрической нити на передней поверхности хрусталика. Можно было изменять положение конденсора и диафрагмы и изменять ось фиксации, но все равно изображение оставалось замутненным, неясным, с искаженными границами. Подобное затруднение было обусловлено тем, что свет не был настроен под самым оптимальным для достижения необходимого эффекта углом и было не всегда возможным определить точную ось, вдоль которой может быть получено ясное, разборчивое изображение. Как и в случае с отражениями от боковых поверхностей глазного яблока, создавалось впечатление, что изображение изменяется без явной на то причины. Однако это было правдой – то, что были углы расположения оси глазного яблока, дававшие более хорошие изображения, нежели остальные, и их невозможно было точно определить. Я работал над освещением по два-три часа, но так и не мог подобрать правильного угла. Бывало и так, что ось оставалась неизменной в течение нескольких дней, позволяя тем самым всегда получать чистые и разборчивые изображения.
Результаты этих экспериментов подтвердили выводы, полученные из предыдущих экспериментов, а именно то, что аккомодация происходит за счет удлинения глазного яблока, а не за счет изменения кривизны хрусталика. Они также поразительным образом подтвердили мои более ранние заключения по поводу состояний, при которых возникают миопия и гиперметропия.[48]
Изображения, сфотографированные с передней поверхности хрусталика, не показали никакого изменения размера или формы во время аккомодации. Как показал телескоп офтальмометра, изображения на задней поверхности хрусталика также остались без изменения, но так как дискуссии по поводу ее поведения во время аккомодации не ведутся, то изображение, отраженное от нее, не было сфотографировано. Изображения с радужки до и во время аккомодации были одинакового размера и формы, как и ожидалось, исходя из характера изображений на поверхности хрусталика. Если форма хрусталика изменяется во время аккомодации, то радужка, которая закрывает его, также должна изменяться.