



Марина Геннадьевна Колбенева
Регрессия как этап развития
2.1. Формирование субъективного опыта как системная дифференциация
• Реализация поведения есть реализация истории его формирования, т. е. активация множества систем, каждая из которых фиксирует этап становления данного поведения.
К наиболее общим законам развития может быть отнесен принцип дифференциации. Его применение позволяет а) отобразить формальные свойства организации и тем самым дать описание разнообразных процессов в общих терминах, б) охарактеризовать динамику процессов, в) связать настоящее и прошлое в поведении индивидов и групп (Werner, 1962, p. VI).
Рядом авторов, находящихся на разных позициях, были приведены аргументы в пользу того, что индивидуальное развитие может быть рассмотрено как нарастающая дифференциация и сложность в соотношении индивида со средой (Александров, 1989, 2011а; Шмальгаузен, 1982; Чуприкова, 1997; Левин, 2001; Сергиенко, 2006; Черниговская, 2013; Lewin, 1937; Werner, Kap lan, 1956; Anokhin et al., 1996; Tononi, Edelman, 1998; Alexandrov, 1999a, b; и др.). Формирование новых систем в процессе индивидуального развития обусловливает прогрессивное увеличение дифференцированности, «подробности» в соотношении организма и среды (подробнее см.: Александров, 1989, 2006, 2011а, б; Чуприкова, 1997).
Упрощая, можно описать рост дифференцированности соотношения со средой следующим образом. Находясь в утробе матери, плод обеспечивает приток материнской крови и, следовательно, питательных веществ и кислорода к плаценте, совершая самые разнообразные движения. После рождения ему необходимо для достижения той же общей «метаболической» цели совершать весьма специализированные движения: дышать, чтобы получать кислород, захватывать сосок материнской груди (или соску), чтобы получать пищу. Затем оказывается, что кроме молока можно получать, например, сок из ложечки, для потребления которого надо совершать питьевые движения. Затем оказывается, что можно еще есть твердую пищу, которую надо жевать. Потом, что можно есть из чашечки, из тарелки, с помощью разных приборов и манипуляций, самую разную пищу (твердую, жидкую, смешанную, овощи, фрукты и т. д.), совершая разные типы движений и ориентируясь на самые разные свойства пищи, определяемые зрительно, по запаху, тактильно, по вкусу. Потом оказывается, что пищу можно получать не только дома, но еще и в школе, в кафе, на улице, в гостях, причем ее получение включает целый ряд специальных подготовительных действий и учет множества факторов, например, наличие свободных мест в кафе и денег.
В основе образования нового элемента субъективного опыта (системы) лежит не «переспециализация» ранее специализированных нейронов, а установление постоянной специализации относительно вновь формируемой системы части ранее «не задействованных», сформированных в раннем онтогенезе нейронов «резерва» или клеток, появившихся при неонейрогенезе (т. е. в ходе образования нейронов во взрослом мозге de novo).
В процессе развития происходит изменение структуры субъективного опыта: новые, более дифференцированные функциональные системы «добавляются» к ранее сформированным системам, не разрушая их, а «наслаиваясь» на них. Однако при формировании новых систем ранее сформированные системы не остаются неизменными. Они претерпевают модификацию за счет процесса «аккомодационной» (приспособительной) реконсолидации (повторной консолидации, в результате которой существующий опыт «приспосабливается» ко вновь формируемому); реконсолидация проявляется в изменении активности, в том числе генетической, ряда нейронов, специализированных в отношении ранее сформированных систем (Александров, 1989, 2001, 2011; Александров и др., 2015; Сварник и др., 2014; Alexandrov et al., 2001; Alexandrov, 2008), поэтому реконсолидационная модификация памяти – никогда не прекращающийся процесс (McKenzie, Eichenbaum, 2011). Таким образом, субъективный опыт индивида представляет собой структуру, образованную системами («слоями») разного «возраста» и разной степени дифференцированности.
Проведенные нами исследования (см.: Швырков, 2006; Александров, 2011б) показали, что осуществление поведения обеспечивается не только посредством реализации новых наиболее дифференцированных систем, сформированных при обучении новому поведению, но и посредством одновременной актуализации множества менее дифференцированных систем, сформированных на предыдущих этапах индивидуального развития. Фактически реализация поведения есть реализация истории его формирования, т. е. множества систем, каждая из которых фиксирует этап становления данного поведения. Обращаясь к упрощенному описанию, данному в начале этого раздела, можно сказать, что при еде в кафе актуализируется не только сравнительно дифференцированный опыт потребления пищи, но и тот менее дифференцированный, что был получен на самых ранних этапах онтогенеза при формировании систем, обеспечивающих достижение пищедобывательных целей.
2.2. Единая концепция сознания и эмоций
• Эмоции и сознание – характеристики разных, одновременно актуализируемых уровней системной организации поведения.
Системы, формирующиеся на самых ранних стадиях индивидуального развития, обеспечивают минимальный уровень дифференциации: хорошо – плохо; приближение (approach) – избегание (withdrawal). Это разделение применимо ко всем живым существам. В единой концепции сознания и эмоций приведены аргументы (см.: Александров Ю. И., 2006; Alexandrov, Sams, 2005) в пользу того, что эмоции преимущественно характеризуют активность систем, сформированных на самых ранних этапах развития («старых» систем), и соотносимы со сравнительно низкодифференцированными уровнями организации поведения.
Представления о том, что при переживании интенсивных эмоций происходит «возврат» к реализации преимущественно «простых» форм поведения, высказывались и ранее. Так, П. Жане указывал на то, что сильная эмоция является «реакцией отката назад или упрощения – знаменует возврат к элементарным и примитивным формам реагирования» (Жане, 1929/2010, с. 92). В своей феноменологической теории эмоций Ж.-П. Сартр утверждал, что «в эмоции сознание деградирует и внезапно преобразует мир причинных связей, в котором мы живем, в магический мир» (Сартр, 1948/1984, с. 134). Он отмечал, что «не имея возможности в состоянии высокого напряжения найти тонкое и точное решение проблемы, мы действуем на самих себя, мы „опускаемся“ и превращаем себя в такое существо, которое способно удовлетвориться грубыми и менее адаптированными решениями (например, разорвать листок, на котором написаны условия задачи)» (Сартр, 1948/1984, с. 125).
Основное положение единой концепции сознания и эмоций состоит в том, что сознание и эмоции являются характеристиками разных, одновременно актуализируемых уровней системной организации поведения, представляющих собой трансформированные этапы развития и соответствующих различным уровням системной дифференциации (высокой и низкой).
При этом сознание и эмоции рассматриваются континуально: они являются различными характеристиками единой системной организации поведения, а не отдельными (дизъюнктивными) психологическими процессами, которые обеспечиваются различными нейрофизиологическими механизмами. Континуальность в единой концепции сознания и эмоций означает, что в развитии нет критического момента появления сознания или исчезновения эмоций. На каждом этапе развития, на каждом уровне системной дифференциации поведение может быть описано с применением обеих характеристик. Однако на каждом уровне соотношение этих характеристик различно. Для каждого данного этапа развития такая характеристика, как эмоции, максимально выражена для наименее дифференцированных систем. Поскольку высокодифференцированные системы, формируясь, не заменяют низкодифференцированные, поведение любого индивида обладает обеими этими характеристиками, выраженность которых зависит от ряда факторов. Одним из этих факторов является относительный вклад в реализацию поведения систем низкой и высокой дифференциации.
Как было отмечено выше, каждый акт поведения есть актуализация систем разного уровня дифференциации, однако соотношение активных высоко- и низкодифференцированных систем, необходимых для успешной реализации поведенческого акта, различается для разных актов. Из единой концепции сознания и эмоций следует, что чем выше доля нейронов низкодифференцированных систем, активных в реализующемся поведении, тем выше интенсивность эмоций. В связи с этим можно предположить, что подавление активности нейронов, принадлежащих к высокодифференцированным системам, должно вести к относительному увеличению вклада систем низкой дифференциации и увеличению интенсивности эмоций как их характеристики.
Глава 2. Регрессия при острой алкогольной интоксикации
1. Мозговые механизмы регрессии
• Регрессия при введении алкоголя имеет в своей основе врéменное угнетение активности нейронов более новых дифференцированных систем – элементов субъективного опыта, т. е. врéменную дедифференциацию.
• Дедифференциация – обратимое относительное увеличение представленности в актуализированном опыте менее дифференцированных систем.
• Увеличение относительного вклада рано сформированных менее дифференцированных систем в обеспечение поведения связано с увеличением интенсивности эмоций.
• Алкоголь сильнее влияет на новый опыт, чем на старый, и на более сложное поведение, чем на простое.
В многочисленных экспериментах, проведенных нами, было показано, что под влиянием алкоголя происходит относительное увеличение вклада реализации систем низкой дифференциации в обеспечение поведения как у человека, так и у животных. Его острое введение (этанол; 1 г/кг) животным вызывает достоверное и обратимое уменьшение числа активных в поведении нейронов мозга, принадлежащих к наиболее новым и дифференцированным системам (Alexandrov et al., 1990а, 1991, 2013; позднее сходные данные были получены и другими авторами; см. ссылки в Alexandrov et al., 2013). Например, в лимбической коре их относительное число уменьшается в 2,5 раза.
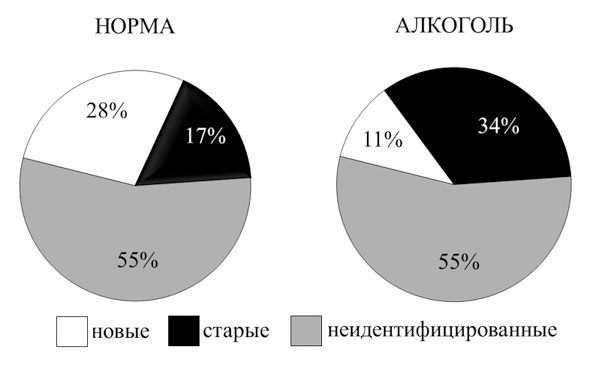
Рис. 1. Процент нейронов разной специализации в лимбической коре кролика в норме (инъекция физиологического раствора) и после введения алкоголя
Относительное число нейронов с неидентифицированной специализацией (н/и; не удается выявить постоянную связь изменения частоты их активности с актами анализируемого поведения) не меняется. В то же время видно уменьшение (с 28 % до 11 %) относительного числа активных нейронов, принадлежащих к наиболее новым (Нов) дифференцированным системам, сформированным на последнем, непосредственно предшествующем регистрации активности нейронов этапе индивидуального развития (обучение животных инструментальному пищедобывательному поведению), и увеличение (с 17 % до 34 %) относительного числа активных нейронов, принадлежащих к системам, сформированным на ранних этапах индивидуального развития («старые»; Ст). При этом достоверно возрастало число ошибок в поведении. Специальные подсчеты (произведенные при учете числа активных нейронов, обнаруженных при движении микроэлектрода сквозь толщу коры) показывают, что отмеченное выше изменение относительного числа нейронов обусловлено достоверным уменьшением абсолютного числа активных нейронов новых систем после введения алкоголя. После завершения действия алкоголя число активных нейронов новых систем возвращается к норме.
Более выраженное угнетающее влияние (как острое, так и хроническое – Alexandrov et al., 2001, 2013) алкоголя на новые сравнительно более дифференцированные системы (согласующееся с принципом Рибо – Джексона – Ribot, 1901) было выявлено нами также при сопоставлении эффектов этанола на медленные электроэнцефалографические потенциалы мозга, регистрируемые у птенцов при актуализации систем, обеспечивающих поведенческие акты, которые последовательно формируются на самых ранних стадиях онтогенеза. Было выявлено, что при введении алкоголя реализация вновь сформированного более сложного (оборонительного) поведения сопровождается более выраженным избирательным угнетением мозговой активности (уменьшением амплитуды ЭЭГ-потенциалов, вызванных значимым сигналом) по сравнению с ранее сформированным (пищедобывательным) поведением (Alexandrov, Alexandrov, 1993).
Мы исследовали влияние острого введения алкоголя на мозговую активность незрелорождающихся птенцов мухоловки-пеструшки через 4–8 дней после их вылупления. Птенцам предъявляли звуки (посылки чистых тонов), связанные с реализацией естественного поведения (2,0 и 5,0 кГц; пищедобывательное и оборонительное поведение, соответственно), а также контрольный звук (тон), не имеющий поведенческой значимости для птенцов данного возраста (3,0 кГц). Регистрировали вызванные ЭЭГ-потенциалы при предъявлении этих звуков на последовательных стадиях формирования и последующего увеличения дифференцированности поведения. После регистрации вызванных потенциалов птенцу давали личинку мучного хрущака, в которую непосредственно перед этим вводили этиловый спирт (96 %) из расчета дозы 1–1,2 г/кг массы тела. Острое введение алкоголя птенцам мухоловки-пеструшки приводило к уменьшению амплитуды слуховых вызванных потенциалов, возникающих в поле L каудального неостриатума в ответ на предъявление звуков, связанных с пищевым и оборонительным поведением, но не контрольного звука (рисунок 2). Анализ динамики амплитудно-временных показателей (коэффициента зрелости) вызванных потенциалов при введении алкоголя позволил сделать вывод о том, что имеет место сдвиг показателей в сторону меньшей зрелости. Подчеркнем, что изменения становились достоверными на 5-й день постэмбрионального онтогенеза, когда у птенцов вместе с раскрыванием глаз появляется новая форма поведения – оборонительное, а ранее сформированное пищевое поведение существенно модифицируется, становится более дифференцированным, формирующимся с учетом ранее не учитываемых деталей среды.

Рис. 2. Усредненный вызванный потенциал 5-дневного птенца на тон 5 кГц до (сплошная линия) и через 10 минут после (пунктирная линия) введения алкоголя. По горизонтали – время, мс (начало эпохи анализа совпадает с началом предъявления тона); по вертикали – амплитуда, мкВ (позитивность вниз). Вертикальные линии на пике N1 – стандартная ошибка значения амплитуды
В цитированных выше работах было показано, что алкоголь угнетает активность нейронов, принадлежащих к наиболее новым системам. В экспериментах с участием добровольцев мы (Alexandrov et al., 1998) проверяли, сохраняется ли повышенная чувствительность к алкоголю, отмеченная для только что образованных систем и для относительно более новых и дифференцированных систем в сравнении с системами, сформированными на более ранних этапах развития. Было сопоставлено влияние острого введения алкоголя (этанол; 1 г/кг) на мозговые механизмы категоризации слов родного (финского) и позже усвоенного (в процессе школьного и университетского обучения) иностранного (английского) языка. Было обнаружено, что острое введение алкоголя в большей степени снижает амплитуду ЭЭГ-потенциалов, связанных со словами иностранного языка, чем потенциалов, связанных со словами родного языка (освоение которого начинается уже в пренатальном периоде). На основе проведенного дисперсионного анализа мы выявили, что этанол угнетает в достоверно большей степени амплитуду раннего негативного ЭЭГ-потенциала N100, связанного с предъявлением английских слов, по сравнению с N100, возникающим при предъявлении финских слов. В таблице 1 сопоставлена выраженность эффекта этанола при категоризации английских и финских слов. Представлено уменьшение амплитуды (в %) в ситуации после введения этанола по сравнению с контрольной ситуацией (без введения этанола).
Видно, что в каждом из отведений угнетающий эффект этанола был более выражен для английских слов. Видно также, что, как и в случае со сравнительно коротколатентным N100, этанол угнетал в большей степени и поздний (латенция более 400 мс) негативный потенциал, связанный с категоризацией английских слов. Поскольку было показано, что использование родного и иностранного языка обеспечивается активностью, по крайней мере, частично неперекрывающихся совокупностей нейронов (см., напр.: Ojemann, 1990), наши данные могут служить аргументом в пользу предположения о том, что введение этанола угнетает преимущественно не только нейроны вновь образованных систем, но и нейроны относительно более новых и дифференцированных систем в сравнении с системами, сформированными на более ранних этапах индивидуального развития.
Таблица 1. Уменьшение (в %) амплитуды N100 и позднего негативного потенциала (латенция более 400 мс) после приема алкоголя по сравнению с контролем (100 %)

Поскольку научение родному языку связано с формированием систем на самых ранних этапах онтогенеза, логично предположить, что актуализация этих систем характеризуется большей эмоциональностью, чем актуализация позднее формирующихся систем, связанных с использованием иностранного языка. Действительно, было обнаружено, что высказывания на родном языке, предъявляемые участникам исследования, воспринимаются ими более эмоционально, чем высказывания на иностранном языке (Harris et al., 2003; Puntoni et al., 2009).
Если алкоголь избирательно блокирует более новые системы, можно ожидать, что его введение не только сильнее угнетает электрические потенциалы головного мозга, связанные с категоризацией слов именно иностранного языка, но и усиливает эмоциональность, так как представленность сравнительно более старых (менее дифференцированных) элементов в наборе актуализированных систем увеличивается. Действительно, в проведенных нами экспериментах при введении алкоголя наблюдалось выраженное усиление эмоциональности – эйфория – и одновременно увеличение числа ошибок при категоризации слов (Alexandrov et al., 1998).
Мы также обнаружили, что острое введение алкоголя приводит к статистически значимому усилению эмоциональности и в ситуации психологического тестирования (Бодунов и др., 1996). Участники исследования проходили двукратное тестирование (один раз в трезвом состоянии и один раз под воздействием алкоголя в дозе 1 мл 96 %-ного этанола на 1 кг веса). Использовали Павловский темпераментальный опросник (Бодунов, Романова, 1993), личностный опросник «Пять факторов» (ОПФ) (Big five, П. Т. Коста, Р. Р. Маккрей, адаптирован М. В. Бодуновым), а также опросник структуры темперамента (ОСТ) (Русалов, 1990). Действие алкоголя было сопряжено с ростом показателя «силы нервной системы» и повышением эмоциональной чувствительности (усилением нейротизма, измеряемого опросником «Пять факторов», и увеличением социальной эмоциональной чувствительности, измеряемой ОСТ).
Полученные нами данные о выраженном подавлении актуализации систем более нового, дифференцированного поведения при остром введении алкоголя согласуются с результатами других авторов, демонстрирующими, что алкоголь сильнее влияет на новый опыт, чем на старый, и на более сложное поведение, чем на простое (Huntley, 1974; Alkana, Malcolm, 1986; Howat et al., 1991; Jääskeläinen et al., 1999; White, 2003).
В качестве признака регрессии при введении алкоголя указывается примитивизация поведения (Salvatore, 1972, 1975). Дополнительным компонентом примитивизации, а также отмеченного угнетения активности более новых систем и увеличения активности более старых и менее дифференцированных систем может быть нарушение контроля ошибок в поведении, физиологическим выражением чего является угнетение ЭЭГ-потенциалов, связанных с функционированием «мозговых механизмов детекции ошибок» (Киреев и др., 2008; Ridderinkhof et al., 2002; Nelson et al., 2011). В свою очередь, с этим эффектом может быть связано возникающее у человека при остром введении алкоголя нарушение учета ошибок при обучении, в том числе в виртуальной среде.
В использованной нами (Безденежных, Александров, 2014) методике тестирования людям предлагали обучиться навигации в водном лабиринте Морриса. Участникам исследования сообщали, что они будут находиться в виртуальном бассейне и их задача заключается в том, чтобы при «плавании» как можно быстрее найти скрытую под водой платформу и выбраться на нее. Люди проходили это тестирование в обычном состоянии и состоянии алкогольного опьянения с интервалом в 2–3 месяца. В работе анализировали общее время, необходимое для нахождения платформы, время остановок для смены направления «плавания», тактику «плавания», а также эффект последовательности, т. е. влияния характеристик предшествующей реализации поведения на характеристики последующей. В результате сравнения динамики обучения ориентированию в виртуальном лабиринте под действием острого введения этанола и в контроле было обнаружено, что время поиска у людей, не принимавших алкоголь, зависело от последовательности предшествующих проб (df 1/20, F = 3,199, p = 0,020 и df 2/20, F = 3,163, p = 0,013). В состоянии же алкогольного опьянения время поиска платформы со стартовой позиции 1 и 2 не зависело от последовательности предшествующих проб (df 3/19, F = 0,336, p = 0,84 и df 4/19, F = 0,311, p = 0,63). Результаты данного эксперимента указывают на то, что у «алкогольной» группы эффект последовательности не выявляется. Можно предположить, что алкоголь, подавляя активность нейронов дифференцированных систем, существенно влияет на детальность, точность процесса прогнозирования, которые связаны с учетом оценки результатов предшествующих действий при планировании следующих действий. «Огрубление» данного процесса оказывается важным фактором, влияющим на формирование памяти.
Неудивительно, что повышенная эмоциональность и пониженный контроль ошибок в поведении обусловливают неадекватно завышенное представление участников эксперимента об эффективности собственного поведения (Conger, 1958; Alexandrov et al., 1998).
Таким образом, полученные результаты указывают на то, что увеличение относительного вклада рано сформированных менее дифференцированных систем в обеспечение поведения связано с увеличением интенсивности эмоций[4]. С этими результатами согласуются и данные, полученные при картировании активности мозга человека в условиях высокоэмоционального состояния – оргазма (Tiihonen et al., 1994; Georgiadis et al., 2006). Было обнаружено, что во время оргазма активность корковых областей, в которых особенно велико число нейронов высокодифференцированных систем (см.: Alexandrov et al., 2000), как правило, достоверно уменьшается.
