



Ричард Докинз
Рассказ предка. Паломничество к истокам жизни
Рассказ Человека работающего
Ричард Лики очень трогательно рассказывает, как его коллега Камойя Кимеу 22 августа 1984 года нашел “мальчика с озера Туркана” (Homo ergaster). Возраст его – 1,5 млн лет, и это древнейший из известных скелетов гоминид, сохранившийся почти полностью. Не менее трогательно Дональд Джохансон описывает обнаружение еще более древнего и поэтому хуже сохранившегося скелета австралопитека, который известен под именем Люси. Достойно внимания и открытие Stw 573 (Little Foot), еще не до конца описанного. Мы не знаем, что позволило “Люси”, Stw 573 и “мальчику с озера Туркана” обрести своего рода бессмертие. Было бы здорово, если бы в свое время то же самое произошло с нами. Как вообще формируются окаменелости?
Горные породы сложены из кристаллов, хотя часто они слишком мелкие, чтобы их можно было разглядеть невооруженным глазом. Кристалл представляет собой гигантскую молекулу, атомы которой располагаются в узлах решетки. Узор решетки повторяется миллиарды раз. Атомы выстраиваются на растущем краю кристалла из жидкости (обычно это вода, в некоторых случаях – сам расплавленный минерал). Форма кристалла и углы пересечения его граней повторяют в большем масштабе строение атомной решетки. Иногда проекция может быть по-настоящему крупной – например в алмазе или аметисте, грани которых выдают трехмерную структуру самоорганизованной атомной решетки. Однако обычно кристаллические единицы, из которых сложены породы, слишком малы – поэтому, в частности, большинство горных пород непрозрачны. К самым важным и распространенным кристаллическим породам относятся кварц (диоксид кремния), полевые шпаты (в основном тоже диоксид кремния, но некоторые из атомов кремния в них заменены на атомы алюминия) и кальцит (карбонат кальция). Гранит – плотно упакованная смесь кварца, полевого шпата и слюды, которая кристаллизуется из магмы. Известняк – это в основном кальцит, песчаник – в основном кварц. В обоих случаях тонкие перемолотые слои затем спрессовываются из отложений песка или ила.
Вулканические породы образуются из остывающей лавы (которая, в свою очередь, является расплавленной породой). Зачастую, как в случае гранита, они бывают кристаллическими. Иногда они могут выглядеть как стекловидная затвердевшая жидкость, а иногда, если сильно повезет, расплавленная лава застывает в “форме для отливки”, например в следе динозавра или пустом черепе. Однако главная польза от вулканических пород для ученых, изучающих историю жизни на земле, заключается в датировании. Мы узнаем из “Рассказа Секвойи”, что лучшие методы датирования применяются лишь для вулканических пород. Сами окаменелости обычно нельзя датировать точно, однако всегда можно поискать поблизости вулканические породы. Тогда мы либо считаем, что окаменелость и порода – ровесники, либо ищем два поддающихся датированию образца вулканической породы, между которыми залегает окаменелость, и устанавливаем верхнюю и нижнюю границы ее возраста. “Бутербродный” метод датирования отчасти рискованный, поскольку труп мог переместиться в другое место из-за наводнения или, например, его могли перетащить гиены или динозавры. К счастью, обычно это видно. Если же нет, приходится прибегать к сопоставлению с общими статистическими закономерностями распределения окаменелостей.
Песчаник, известняк и другие осадочные породы сложены из крошечных частиц, выветренных или вымытых водой из более древних пород или других твердых материалов, например раковин моллюсков. Эти частицы переносятся в составе взвеси – например песка, ила либо пыли, – постепенно спрессовываясь в новые слои породы. Большая доля окаменелостей обнаруживается в осадочных породах.
Осадочные породы устроены так, что вещества, их составляющие, подвергаются непрерывной рециркуляции. Древние горы, например Северо-Шотландское нагорье, подтачивают ветер и вода. Образовавшиеся частицы оседают и могут снова подняться в другом месте, например в молодых горах вроде Альп, и тогда цикл повторится. Учитывая этот круговорот, мы должны понимать, что невозможно сделать непрерывную палеонтологическую летопись, которая заполняла бы все пробелы в эволюции. Мы не всегда можем найти окаменелости, и это прямое следствие процесса образования осадочных пород. Наоборот, было бы подозрительно, если бы в летописи не было пробелов. Древние породы и хранящиеся в них окаменелости постоянно разрушаются.
Нередко окаменелости формируются в результате проникновения насыщенной минералами воды в ткани захороненного организма. При жизни кости пористые, и на то есть “технические” и “экономические” причины. Когда вода просачивается через поры мертвой кости, на ней медленно откладываются минералы. Впрочем, вспомните, как быстро в чайнике образуется накипь. (Однажды в Австралии я нашел на побережье крышку от бутылки, заключенную в камень.) Но обычно процесс все-таки медленный. Так или иначе, образуется окаменелость в форме исходной кости, и эту окаменелость мы находим миллионы лет спустя, даже если (не всегда) от кости не осталось ни единого первоначального атома. Окаменелый лес в национальном парке Петрифайд-Форест (Аризона) состоит из деревьев, ткани которых медленно заменялись кварцем и другими минералами из грунтовых вод. Эти деревья мертвы уже 200 млн лет, они окаменели, но мы до сих пор можем рассмотреть их клетки.
Иногда исходный организм или его часть оставляет естественную форму или отпечаток. Сам организм впоследствии исчезает из этой формы или разлагается. Я с удовольствием вспоминаю два прекрасных дня в 1987 году в Техасе, в долине реки Пелекси, где я рассматривал следы динозавра на гладком известняке и даже ставил в них ноги. Местная легенда гласит, что некоторые из этих следов принадлежат гигантскому человеку – современнику наследившего там динозавра. В соседнем городе Глен-Роуз бурно расцвел кустарный промысел: там весьма неуклюже изготавливают гигантские отпечатки ног человека в цементе – для продажи наивным креационистам, верящим, что “в те дни на земле жили исполины” (Бытие 6:4). Что касается настоящих следов, то их историю реконструировали. Трехпалые следы бесспорно принадлежат динозаврам. А следы, напоминающие след человеческой ноги, принадлежат динозаврам, которые ступали на пятки, а не на носки. Кроме того, через края следа, скорее всего, перетекала жидкая грязь, делая нечеткими отпечатки боковых пальцев. Еще больший интерес представляет находка в Лаэтоли (Танзания). Там обнаружились следы трех настоящих гоминид, вероятно Australopithecus afarensis, которые 3,6 млн лет назад прошли по еще свежему вулканическому пеплу. Кто из нас не задумается над тем, кем приходились эти люди друг другу? Держались ли они за руки? Может, даже разговаривали? И какое забытое ныне дело было у них в то плиоценовое утро?
Иногда (вспомните разговор о вулканической лаве) форма может заполняться каким-нибудь материалом, впоследствии затвердевающим и образующим слепок животного или части его тела. Я пишу эти строки в саду, на столе, столешница которого (квадратная двухметровая плита пятнадцатисантиметровой толщины) представляет собой кусок пурбекского известняка, возраст которого, возможно, составляет 150 млн лет[7]. Наряду со множеством окаменелых раковин моллюсков, на нижней поверхности этой плиты якобы (по словам одного известного эксцентричного скульптора, который добыл ее для меня) имеется след динозавра. Правда, след этот объемный, выступающий над поверхностью. Исходный отпечаток (при условии, что он настоящий – на мой взгляд, он не выглядит убедительно) должен был служить формой, в которой накапливались осадочные породы. Сама форма после этого исчезла. Большей частью знаний о мозге древних людей мы обязаны дошедшим до нас “отливкам” черепов. Нередко на них неожиданно четко отпечатывается поверхность мозга.
Иногда фоссилизируются и мягкие ткани животных, хотя не так часто, как раковины, кости или зубы. Самые известные места таких находок – сланцы Берджес в Канадских Скалистых горах и немного более древний палеонтологический парк Чэнцзян в Южном Китае (туда мы вернемся в “Рассказе Онихофоры”). И там, и там наряду с обычными “твердыми” ископаемыми найдены окаменелые остатки червей и других мягких беспозвоночных беззубых животных, которые относятся к кембрию (более 500 млн лет назад). Нам очень повезло, что существуют Чэнцзян и Берджес. Более того, как я уже говорил, нам очень повезло, что у нас вообще есть ископаемые. 90 % видов никогда не встретятся нам в виде ископаемых. Если это число справедливо для одного вида, подумайте, как мало людей могло стать окаменелостями! Согласно одной оценке, для позвоночного вероятность такого исхода составляет 1: 1000000. Мне это соотношение кажется даже преувеличенным, ну а для животных, не имеющих твердых органов, оно и того меньше.
Человек умелый
Если отступить от времени Homo ergaster еще на 1 млн лет в прошлое, у нас не останется сомнений в том, на каком континенте следует искать свои корни. Ученые (включая сторонников “мультирегионального” происхождения человека) согласны в том, что это – Африка. Самые убедительные находки костей указанного возраста обычно относят к H. habilis. Некоторые авторитетные ученые выделяют еще один современный ему вид – H. rudolfensis. Другие отождествляют его с кениапитеком, описанным Лики и его коллегами в 2001 году. Есть специалисты, которые вообще предпочитают не давать этим окаменелостям видовые названия, а просто относят их всех к “раннему Homo”. У меня нет по этому вопросу четкой позиции. Мне важнее люди из плоти и крови, и я буду называть их “умелыми”. Ископаемые остатки людей умелых старше, чем остатки людей работающих, и их сохранилось меньше. Череп самой хорошей сохранности имеет номер KNM-ER 1470. Его хозяин жил около 1,9 млн лет назад.
H. habilis столь же сильно отличался от человека работающего, как человек работающий – от нас самих. Существуют также промежуточные формы, которые довольно трудно классифицировать. Череп H. habilis был не таким прочным, как у человека работающего, а надбровные дуги не были так сильно выражены. В этом отношении H. habilis были похожи на нас, и это не удивительно. Прочность черепа и надбровные дуги, судя по всему, относятся к признакам, которые (например, волосы) гоминиды могут терять или приобретать по малейшему эволюционному поводу. Начиная с H. habilis, наш мозг увеличивался. Точнее, его размеры начали превышать средние размеры мозга других человекообразных обезьян – хотя и у них мозг уже был довольно крупным. Именно поэтому человека умелого относят к роду Homo. Для многих палеонтологов крупный мозг – отличительный признак нашего рода. H. habilis, объем мозга которого превысил 750 см3, перешел Рубикон и стал человеком.
Я не люблю “рубиконы”, “рубежи” и “промежутки”. В частности, нет оснований полагать, что ранний H. habilis отличался от предшественника сильнее, чем от преемника. Эта мысль может показаться здравой, потому что предшественник носит другое родовое название (австралопитек), а преемник (H. ergaster) – просто “другой Homo”. Конечно, если мы рассматриваем современные виды, мы ждем, что представители разных родов будут меньше похожи, чем представители разных видов в пределах одного рода. Однако в случае окаменелостей это не работает, потому что там имеется непрерывный ряд поколений. На границе между любым ископаемым видом и его непосредственным предшественником всегда найдутся особи, о которых можно спорить бесконечно. Потому что, если воспользоваться методом доведения до абсурда, окажется, что родители, принадлежащие к одному виду, породили потомство, принадлежащее уже к другому виду. Еще более нелепо предположение о том, что ребенок рода Homo появился у родителей совершенно другого рода – австралопитеков. Таким образом, в этой области науки об эволюции правила таксономии не работают.
Отказ от названий позволяет рассуждать о том, почему мозг внезапно начал расти. Как оценить увеличение мозга гоминид и построить график зависимости среднего размера мозга от геологического времени? С единицами времени проблем не возникает: это должны быть миллионы лет. С размером мозга сложнее. Ископаемые черепа и слепки позволяют оценивать размер мозга в кубических сантиметрах, которые потом довольно легко перевести в граммы. Однако нам не обязательно нужен абсолютный размер мозга. У слона мозг крупнее нашего, и тем не менее мы с полным правом считаем себя умнее. Мозг тираннозавра был ненамного меньше нашего, однако это не мешает считать динозавров глупыми: относительный размер нашего мозга больше.
Можно вносить поправку на абсолютный размер и выражать размер мозга как функцию от размера, который “должен” иметь мозг при данном размере тела. Эта тема достойна отдельного рассказа.
Рассказ Человека умелого
Итак, мы хотим оценить размер мозга животного – например Homo habilis: больше он или меньше, чем “должен быть” при данном размере тела? Мы допускаем (я – не очень охотно, но так и быть), что у крупных животных мозг должен быть крупным, а у малых – маленьким. Мы, тем не менее, желаем знать, бывают ли одни виды “мозговитее” других. Как рассчитать поправку на размер тела? Нам нужно сформулировать разумный принцип, согласно которому мы сможем вычислить прогнозируемый размер мозга животного на основе размера его тела. И тогда мы сможем сопоставить реальный размер мозга с прогнозируемым.
Эти вопросы могут относиться к любой другой части тела. Бывает ли так, что сердца, почки или лопатки у некоторых животных крупнее (или мельче), чем должны быть? Если да, то можно предположить, что их образ жизни требует специального размера сердца (почки, лопатки и так далее). Нам известен общий размер его тела. При этом не забывайте, что “должен быть” в данном случае не означает “необходим по функциональным причинам”. Это означает “прогнозируемый размер, полученный на основе сравнения с похожими животными”. Поскольку это “Рассказ Человека умелого”, а самая замечательная черта H. habilis – его мозг, то обсуждать этот вопрос мы будем на примере мозга. Но это не помешает нам получить более общие выводы.
Начнем с диаграммы рассеяния массы мозга относительно массы тела для большого количества видов. Каждый символ на графике (его построил мой коллега, выдающийся антрополог Роберт Мартин) обозначает один современный вид млекопитающих – всего их на графике 309, и расположены они от меньшего к большему. (H. sapiens – значок со стрелкой, а значок рядом – это дельфин.) Жирная линия, соединяющая точки, представляет собой прямую, которая, согласно статистическим расчетам, максимально приближена ко всем точкам.

Логарифмическое соотношение массы головного мозга и тела у различных видов плацентарных млекопитающих. Закрашенные треугольники обозначают приматов. Стрелкой обозначен человек. Martin [185].
Небольшое затруднение, смысл которого сейчас станет понятен, заключается в том, что обе шкалы графика лучше делать логарифмическими. Именно так построен этот график. Здесь видна зависимость логарифма массы мозга животного от логарифма массы его тела. Логарифмическия шкала означает, что шаги, отложенные по горизонтальной (или вертикальной) оси, представляют собой значения, умноженные на некоторое число, например на десять, а не просто последовательный ряд значений. Логарифм с основанием десять удобен тем, что мы можем рассматривать его как количество нулей в числе. Если для того, чтобы получить массу слона, нужно умножить массу мыши на миллион, то это значит, что нужно прибавить к массе мыши шесть нулей, то есть нужно добавить шесть к логарифму первого, чтобы получить логарифм второго. На полпути между ними на логарифмической шкале – в трех нулях – находится животное, которое весит в тысячу раз больше мыши и в тысячу раз меньше слона: например человек. Круглые числа, например тысяча и миллион, легче для понимания. “Три с половиной нуля” лежат где-то между тысячей и десятью тысячами. Заметьте, что “на полпути” по отношению к нулям – это не то же самое, что “на полпути между” по отношению к граммам. В нашем случае это делается автоматически благодаря подсчету логарифмов чисел. Логарифмические величины применяют для интерпретаций простых арифметических величин.
Есть по меньшей мере три убедительных причины для использования логарифмической шкалы. Во-первых, это позволяет уместить на одном графике малую бурозубку, лошадь и синего кита без необходимости тратить тонны бумаги. Во-вторых, это позволяет работать с мультипликативными признаками. Мы не просто хотим узнать, что наш мозг крупнее, чем полагается при данном размере тела. Интересно узнать, что наш мозг, скажем, вшестеро больше, чем должен быть. Такие мультипликативные свойства можно увидеть непосредственно на графике. Для этого и нужны логарифмические шкалы. Третья причина труднее для понимания. Во-первых, на таких шкалах облако точек можно разместить вдоль прямой, а не кривой. Но дело не только в этом.
Представьте, что некий объект – сферу, куб или мозг – вы десятикратно увеличиваете, причем его форма должна остаться прежней. В случае сферы это значит, что ее диаметр увеличился в десять раз. В случае куба или мозга это значит, что в десять раз увеличились ширина, высота и длина. А что происходит с объемом? Он увеличится не в десять, а в тысячу раз. Можно сказать, что объем пропорционален одной трети длины и ее логарифму, умноженному на три.
Все это можно проделать не только с объемом, но и с площадью. Однако площадь увеличивается пропорционально длине, возведенной во вторую степень, а не в третью. (Не зря вторую степень называют квадратом, а третью – кубом.) Объем кусочка сахара определяет его количество и цену. Однако скорость растворения зависит от площади поверхности (это не так-то просто высчитать, потому что по мере растворения кусочка площадь его поверхности уменьшается медленнее, чем объем). Если мы равномерно увеличиваем объект, удваивая его длину (ширину и так далее), площадь поверхности умножается вдвое: 2 х 2 = 4. Если увеличить длину в 10 раз, площадь поверхности надо умножить в десять раз: 10 х 10 = 100 (то есть прибавить два нуля). Логарифм площади поверхности увеличивается пропорционально двойному логарифму длины, а логарифм объема – пропорционально тройному логарифму длины. Двухсантиметровый кусочек сахара будет содержать в восемь раз больше сахара, чем односантиметровый, но растворяться в чае он будет лишь вчетверо быстрее (по крайней мере сначала), потому что это зависит от площади поверхности, которая реагирует с чаем.
Теперь возьмем много кусочков сахара разного размера и построим диаграмму, где на горизонтальной оси отложена масса (она пропорциональна объему), а на вертикальной – начальная скорость растворения кусочка (которую мы полагаем пропорциональной площади). На нелогарифмическом графике точки расположены вдоль кривой. Эту линию трудно интерпретировать. Но если мы отложим на графике логарифм массы и логарифм начальной скорости растворения, картина получится информативнее. На каждое троекратное увеличение логарифма массы будет приходиться двукратное увеличение логарифма площади поверхности. В логарифмическом масштабе точки не расположены вдоль кривой: они распределятся вдоль прямой линии. Более того, угол наклона этой прямой составит два к трем, то есть на каждые два шага по оси площади будет приходиться три шага по оси объема. На каждое двукратное увеличение логарифма площади будет приходиться троекратное увеличение логарифма объема. Угол наклона прямой на двойном логарифмическом графике может быть и другим. Графики подобного рода наглядны, потому что угол наклона прямой позволяет почувствовать, как взаимодействуют объем и площадь. А объем, площадь и сложные отношения между ними чрезвычайно важны для понимания того, как устроены организмы и их части.
Я не так уж силен в математике, но и меня очаровали эти расчеты. Еще сильнее меня восхищает, что этот же принцип годится для любых других фигур – не обязательно куба и сферы, но и для сложных: животных и органов животных, например почек и мозга. Для этого нужно, чтобы изменение размера представляло собой пропорциональное увеличение или уменьшение без изменения формы. Это дает нечто вроде нулевой гипотезы, исходя из которой мы можем оценивать реальные размеры. Так, если тело одного животного десятикратно длиннее тела второго, то его масса будет больше в тысячу раз, но лишь при условии, что у них одинаковая форма. Однако форма тела, судя по всему, закономерно эволюционировала по мере движения от маленьких животных к большим, и сейчас мы увидим, почему.
Форма тела крупных и мелких животных неизбежно различается – хотя бы из-за соотношения площади и объема. Если бы мы сделали из землеройки слона, пропорционально ее увеличив и сохранив форму, она бы не выжила. Она стала бы в миллион раз тяжелее, а это породило бы множество проблем. Одни связаны с объемом (массой). Другие – с площадью поверхности. Третьи – с соотношением этих двух величин или другими факторами. Точно так же, как скорость растворения куска сахара зависит от площади его поверхности, у животного скорость теплоотдачи или испарения воды через кожу пропорциональна площади поверхности его тела. Однако скорость теплопродукции, судя по всему, сильнее зависит от числа клеток, которое, в свою очередь, является функцией от объема.
Землеройка размером со слона сохранила бы веретенообразные ножки, которые быстро подломились бы. Ее мышцы были бы слишком тонкими, ведь сила мышцы пропорциональна не ее объему, а площади поперечного сечения. Мышечное движение – суммарное движение миллионов крошечных волокон. Число волокон, которые можно “упаковать” в мышцу, зависит от площади ее поперечного сечения (линейный размер в квадрате). Но работа, которую должна выполнять мышца – например поддержка слона, – пропорциональна массе слона (линейный размер в кубе). Поэтому слону для поддержания тела необходимо пропорционально больше мышечных волокон, чем землеройке. Соответственно, поперечное сечение мышц слона должно быть большим, чем можно ожидать при простом увеличении масштаба. То же касается объема мышц. По разным причинам это правило относится и к костям. Именно поэтому у слонов и других крупных животных ноги толстые, как стволы деревьев.
Допустим, что животное размером со слона стократно крупнее землеройки. Если форма тела у них одинакова, то площадь поверхности кожи первого животного будет в 10 тыс. раз больше, чем у землеройки, а объем и масса тела – в миллион раз больше. И если сенсорные клетки будут распределены на коже таким же образом, у слона их будет в 10 тыс. раз больше – а значит, обслуживающая их область мозга должна пропорционально увеличиться. Всего у первого животного в миллион раз больше клеток, чем у землеройки, и ко всем им должны подходить капилляры. Сколько километров кровеносных сосудов у крупного животного? Посчитать это довольно сложно, мы займемся этим после. Пока достаточно понимания, что в подобных случаях всегда нужно учитывать правила соотношения объема и площади поверхности. И логарифмический график – это отличный способ интуитивного понимания таких вещей. Основной вывод таков: по мере того, как животные в процессе эволюции становятся крупнее или мельче, форма их тела предсказуемо изменяется.
К этому нас привели размышления о размере мозга. Нельзя сравнивать наш мозг с мозгом H. habilis, Australopithecus или любого другого вида, не делая поправку на размер тела. Для этого нам нужен некий индекс размера мозга, учитывающий поправку на размер тела. Причем мы не можем просто разделить размер мозга на размер тела, хотя уже это было бы лучше сравнения абсолютных размеров мозга. Лучший способ – это использование логарифмических графиков. Нужно построить на графике зависимость логарифма массы мозга от логарифма массы тела для животных разного размера. Отметки, скорее всего, будут располагаться на прямой – примерно так, как на графике выше. Если угол наклона прямой составит 1/1 (то есть размер мозга строго пропорционален размеру тела), это означает, что каждая клетка мозга может обслуживать определенное число клеток тела. Угол наклона 2/3 говорил бы, что мозг подобен костям и мышцам: для определенного объема тела (или количества клеток тела) необходима определенная площадь поверхности мозга. Так каков же угол наклона этой прямой?
Он находится примерно между 1/1 и 2/3. Лучше всего он соответствует 3/4. Почему? Это отдельная история, которую расскажет цветная капуста. (Мозг немного похож на цветную капусту, не так ли?)
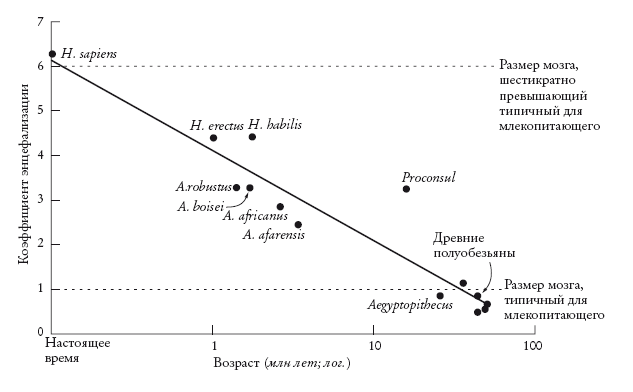
График изменения во времени коэффициента энцефализации (EQ) для различных ископаемых. Время в миллионах лет отложено по логарифмической шкале. Результаты представлены с поправкой на угол наклона 3/4.
Но пока я скажу лишь, что угол наклона 3/4 характерен не только для мозга, но возникает у всех видов живых существ, включая растения – например цветную капусту. В “Рассказе Цветной капусты” мы ознакомимся с интуитивно понятным объяснением этого факта. Что касается размера мозга, то именно 3/4 мы будем использовать, говоря о “прогнозируемом” соотношении, о котором шла речь выше.
Хотя точки на графике группируются вокруг “прогнозируемой” прямой с углом наклона 3/4, с ней совпадают не все точки. Если точка на графике лежит выше прямой, можно сказать, что у животного “большой” мозг – то есть он крупнее, чем “прогнозируемый” при данном размере тела. Животные, у которых мозг меньше “прогнозируемого”, располагаются ниже прямой. Расстояние от точки до прямой говорит о том, насколько мозг животного больше или меньше “прогнозируемого”. Так, точка, лежащая непосредственно на прямой, обозначает животное, размер мозга которого точно равен прогнозируемому.
Когда выполняется это условие? Если животное является типичным представителем группы, для которой строился график. Допустим, график построен для выборки наземных позвоночных от геккона до слона. Если на графике млекопитающие расположены выше прямой (а рептилии – ниже), то это означает, что у млекопитающих мозг крупнее, чем “должен быть” у типичного позвоночного животного. Если теперь мы построим прямую для типичной выборки млекопитающих, она окажется параллельна прямой, построенной для позвоночных. Угол наклона такой же (3/4), однако абсолютная высота будет больше. Прямая, построенная для типичной выборки приматов (высших приматов), еще выше, но угол ее наклона также 3/4. И, наконец, выше всех лежит прямая для H. sapiens.
Мозг человека слишком велик даже по стандартам приматов. А средний мозг примата слишком крупный по стандартам млекопитающих. И, если уж на то пошло, мозг среднего млекопитающего слишком велик для позвоночных животных. Это можно изобразить так: разброс точек для позвоночных шире, чем разброс точек для млекопитающих. В свою очередь, разброс точек для млекопитающих шире, чем разброс для приматов. Или, например, облако точек, обозначающее неполнозубых (отряд южноамериканских млекопитающих, включающий ленивцев, муравьедов и броненосцев), на графике млекопитающих окажется ниже среднего значения.
Гарри Джерисон, стоявший у истоков работ по изучению мозга ископаемых животных, предложил использовать коэффициент энцефализации (Encephalisation Quotient, EQ). Он позволяет оценить отношение действительного размера мозга животного к прогнозируемому размеру мозга при данном размере тела. При этом животное должно принадлежать к многочисленной группе, например к позвоночным или млекопитающим. Это важно: чтобы проводить сравнения, нам нужно определить крупную группу, которая будет использоваться как линия отсчета. Коэффициент EQ для вида – это расстояние от соответствующей точки до средней линии определенной нами крупной группы. Джерисон, правда, думал, что угол наклона этой линии составляет 2/3, тогда как современные ученые сходятся на 3/4. Поэтому оценки EQ, сделанные Джерисоном, нуждаются в некоторых поправках – на это указывал Роберт Мартин. После этого выяснилось, что мозг современного человека примерно вшестеро больше, чем “должен быть” у млекопитающего такого размера. Заметьте, что значение EQ будет еще выше, если его рассчитать применительно ко всем позвоночным, а не только к млекопитающим. И, соответственно, он будет меньше, если рассчитать его для приматов.
Мозг современного шимпанзе примерно вдвое крупнее мозга типичного млекопитающего. Такого же размера мозг был у австралопитеков, H. habilis и H. erectus. Последние два вида, возможно, являются промежуточными между австралопитеками и нами. Размер мозга у них промежуточный. У обоих EQ составляет около 4, то есть их мозг примерно вчетверо больше, чем “должен быть” у млекопитающих такого размера.
На графике выше показаны значения EQ ископаемых приматов и питекантропов в зависимости от времени их существования. С некоторыми оговорками можно считать, что график иллюстрирует уменьшение мозга по мере движения в прошлое. В верхней части графика помещается современный H. sapiens с EQ = 6. Это значит, что наш мозг вшестеро тяжелее, чем у типичного млекопитающего нашего размера. В нижней части графика – ископаемые виды. Кто-то из них, возможно, представляет собой сопредка № 5 – нашего общего предка с обезьянами Старого Света. Их EQ оценивают около 1 – то есть мозг у них был как раз такой, какой полагается типичному млекопитающему их размера. И, наконец, в средней части графика находятся различные виды Australopithecus и Homo, которые вполне могли быть близкими родственниками наших предков. Прямая на графике, повторюсь, проведена так, что расстояние от нее до всех точек минимально.
Это верно с оговорками. Коэффициент EQ подсчитан на основе двух величин: массы мозга и массы тела. В случае ископаемых эти величины оценивают по дошедшим до нас фрагментам. Величина погрешности при этом может быть огромна, особенно при оценке массы тела. Точка на графике, которая относится к H. habilis, показывает, что он был “мозговитей”, чем H. erectus. Я этому не верю. Абсолютный размер мозга H. erectus бесспорно больше. И тот факт, что у H. habilis индекс EQ выше, объясняется предположительно тем, что масса тела у него была меньше. Чтобы почувствовать, насколько велика погрешность, вспомните, как сильно варьирует масса тела у современных людей. Коэффициент EQ чрезвычайно чувствителен к ошибкам при измерении массы тела. Ведь в формуле EQ массу тела, как вы помните, возводят в степень. Таким образом, разброс точек вдоль прямой во многом отражает просто изменчивость массы тела. С другой стороны, тенденция, на которую указывает прямая на графике, судя по всему, вполне реальна. Методы, о которых мы говорим, и особенно оценка EQ на графике, все-таки убеждают, что самое важное событие последних 3 млн лет эволюции – это увеличение мозга у приматов. И возникает вопрос: почему это произошло? Как давление отбора привело к увеличению мозга?




