



Ричард Докинз
Рассказ предка. Паломничество к истокам жизни
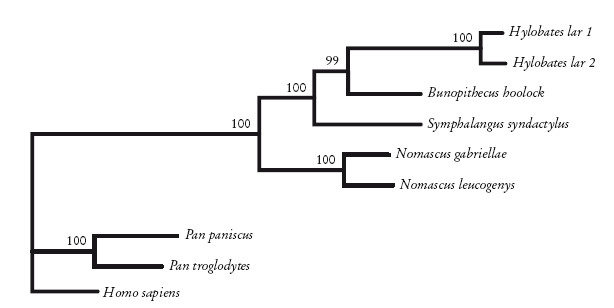
Кладограмма гиббонов, построенная с помощью метода максимального правдоподобия по данным ДНК. Roos and Geissmann [246].
Видообразование у гиббонов произошло сравнительно недавно. Однако если изучать все более удаленные виды, которые будут разделены все более длинными ветвями, в конце концов даже изощренные методы Байеса и максимального правдоподобия откажутся нам служить. В определенный момент недопустимо большая доля сходств окажется случайной. Когда это происходит с ДНК, говорят, что наступило насыщение. И тогда ни один метод не поможет реконструировать схему родственных отношений: действие времени заглушает “филогенетический” сигнал. Особенно остро этот вопрос стоит в отношении нейтральных мутаций ДНК. Давление естественного отбора не позволяет генам сбиваться с пути, удерживая их в узком диапазоне. В некоторых случаях самые важные функциональные гены могут оставаться практически неизменными сотни миллионов лет. Однако для псевдогена, с которого никогда ничего не считывается, таких промежутков времени достаточно для безнадежно сильного насыщения. В таких случаях нам приходится искать другие данные. Одна из самых перспективных идей – использование редких геномных мутаций, о которых я упоминал. Эти изменения затрагивают значительные участки ДНК, а не одну “букву”. Поскольку такие перестройки редки и, как правило, уникальны, проблема случайного сходства не возникает. Эти мутации могут выявлять неожиданные родственные связи. Мы убедимся в этом, когда к толпе пилигримов присоединятся гиппопотамы. (Вот увидите, они расскажут удивительные вещи!)
А теперь обобщим то, что узнали из “Рассказа Митохондриальной Евы” и “Рассказа Неандертальца”. Cчитается, что для группы видов должно существовать лишь одно эволюционное древо. Однако из “Рассказа Митохондриальной Евы” видно, что на основе разных участков ДНК (а также для разных признаков или разных частей тела) можно построить разные деревья. Мне кажется, эта проблема заложена в самой идее филогенетических деревьев видов. Ведь вид представляет собой сложную мозаику фрагментов ДНК, полученных из разных источников. Мы увидели, что каждый ген, да и каждая “буква” ДНК, эволюционирует независимо. Для каждого фрагмента ДНК и каждого признака организма можно построить свое эволюционное древо.
С доказательствами этого мы сталкиваемся каждый день – и поэтому их не замечаем. Если предьявить марсианину гениталии мужчины, женщины и самца гиббона, пришелец, не колеблясь, решит, что наиболее близким родством связаны два самца. И правда: ген, определяющий мужской пол (SRY), никогда не бывал в теле женщины – а если и бывал, то задолго до того, как мы разошлись с гиббонами. Морфологи традиционно делают исключение для половых признаков, избегая “бессмысленных” классификаций. Однако такого рода проблемы встречаются на каждом шагу. Мы столкнулись с этим в “Рассказе Митохондриальной Евы”, когда говорили о группе крови ABo. Если рассматривать гены группы крови, окажется, что мой ген группы крови B сближает меня с шимпанзе с группой крови B, а не A. Все это касается не только генов, определяющих пол, или генов группы крови. Нет, при определенных обстоятельствах эта проблема затрагивает абсолютно все гены и признаки. Большинство молекулярных и морфологических признаков указывает на то, что шимпанзе – наш ближайший родственник. Однако меньшая доля признаков указывает на то, что наш ближайший родственник – горилла, или что шимпанзе ближе всего к гориллам, а не к человеку.
Не удивляйтесь! Популяция, предковая для всех трех видов, должна быть очень изменчивой, и у каждого гена в популяции должно быть несколько вариантов. Каждый из вариантов передается по своей линии. Вполне возможно, например, что человек и горилла получили некий ген от одной линии, а шимпанзе – от другой. После этого нужно только, чтобы разошедшиеся в древности генетические линии тянулись непрерывно до точки расхождения человека и шимпанзе. И получится, что человек произошел от одной линии, а шимпанзе – от другой[16].
Приходится признать, что одно древо не описывает весь эволюционный сюжет. Ничто не мешает нам продолжать строить деревья для видов, однако нужно помнить, что эти деревья представляют не более чем обобщение множества генных деревьев. Интерпретировать деревья можно двумя способами. Первый – традиционная генеалогическая интерпретация. Один вид является ближайшим родственником другого, если из всех рассмотренных видов именно с ним его связывает самый поздний общий предок. Второй способ интерпретации, мне кажется, только предстоит освоить. Согласно этому подходу, построенное для группы видов древо отражает родственные отношения большей части генов. То есть древо показывает результаты, за которые гены высказались “большинством голосов”.
Мне больше нравится идея голосования генов. Поэтому, когда я говорю о родстве видов, его нужно понимать именно так. Все филогенетические деревья, которые я здесь обсуждаю – касаются ли они животных, растений, грибов или бактерий, – нужно рассматривать как схемы, отражающие идеи “генного большинства”.

Узконосые обезьяны. Это общепризнанное филогенетическое древо, построенное примерно для ста видов обезьян Старого Света. (Кружки на концах ветвей указывают на количество видов в каждой группе: отсутствие кружка означает 1–9 известных видов, небольшой кружок соответствует 10–99 видам, круг побольше – 100–999 и т. д. Каждая из представленных здесь четырех групп объединяет 10–99 видов.)
На рис. (слева направо): мандрил (Mandrillus sphinx), краснохвостая мартышка (Cercopithecus ascanius), носач (Nasalis larvatus), ангольский чернобелый колобус (Colobus angolensis).
Рандеву № 5
Обезьяны Старого Света
Приближаясь к рандеву № 5 и готовясь к встрече с сопредком № 5 (нашим прародителем приблизительно в полуторамиллионном поколении), мы пересекаем важный (хотя и произвольно выбранный) рубеж. Впервые с начала путешествия мы вступаем в другой геологический период, палеоген, и оставляем неоген. В следующий раз, пересекая границу, мы окажемся в населенном динозаврами мире – в меловом периоде. Рандеву № 5 проходит около 25 млн лет назад, в олигоценовую эпоху палеогена. Это последняя остановка на дороге в прошлое, во время которой климат и растительность покажутся нам знакомыми. Дальше мы не увидим открытых поросших травой пространств, типичных для неогена, или мигрирующих стад травоядных. Около 25 млн лет назад Африка была полностью изолирована. Даже от ближайшей к ней Испании ее отделял пролив, по ширине равный сегодняшнему Мозамбикскому проливу. На этом огромном острове – Африке – наша команда пополнится новыми участниками, весьма ловкими и сообразительными. Встречайте обезьян Старого Света – первых наших хвостатых спутников.
Сейчас насчитывается почти сто видов обезьян Старого Света. Некоторые когда-то эмигрировали и живут в Азии (см. “Рассказ Орангутана”). Их делят на две крупные группы. Первую составляют африканские колобусы и азиатские лангуры и носачи, вторую – азиатские макаки плюс африканские павианы, мартышки и так далее.
Последний общий предок всех живущих ныне обезьян Старого Света жил примерно на 11 млн лет позднее сопредка № 5, то есть около 14 млн лет назад. Наиболее репрезентативный ископаемый род того времени – викториапитек (Victoriapithecus), от скелета которого сохранилось более тысячи фрагментов, включая отменный череп с острова Мабоко на озере Виктория. Итак, 14 млн лет назад обезьяны Старого Света приветствуют своего предка. Может быть, это сам викториапитек, а может, кто-то похожий на него. Затем пилигримы отправляются в прошлое и на отметке 25 млн лет присоединяются к человекообразным обезьянам и сопредку № 5.
На кого был похож сопредок № 5? Возможно, он напоминал египтопитеков (Aegyptopithecus), которые жили 7 млн лет ранее. Пытаясь реконструировать облик сопредка № 5, применим эмпирическое правило. С высокой вероятностью сопредок № 5 имел признаки, общие для его потомков – узконосых обезьян (Catarrhina), к которым относят человекообразных обезьян и обезьян Старого Света. Так, у сопредка № 5, скорее всего, были узкие, направленные вниз ноздри – в противоположность ноздрям широконосых обезьян Нового Света (Platyrrhina). Для самок, скорее всего, был характерен настоящий менструальный цикл (ежемесячная овуляция), который наблюдается у человекообразных обезьян и обезьян Старого Света, но которого нет у обезьян Нового Света. Вероятно, у сопредка № 5 слуховой проход был обрамлен трубкой барабанной кости, поддерживающей барабанную перепонку, тогда как у обезьян Нового Света барабанная перепонка поддерживается кольцом, и трубки у них нет.
Был ли у сопредка № 5 хвост? Скорее всего, да. Поскольку отсутствие хвоста – это самый наглядный признак человекообразных обезьян, очень хочется сделать вывод, что разделение двух ветвей 25 млн лет назад совпадает со временем утраты хвоста. На самом деле сопредок № 5 был, по-видимому, хвостатым, как и почти все млекопитающие, а сопредок № 4 – бесхвостым, как и все его потомки, современные человекообразные обезьяны. Но в какой момент между сопредками №№ 4 и 5 утрачен хвост, мы не знаем. Это, впрочем, не так важно: мы же не думаем, что обычная обезьяна, утратив хвост, вдруг стала человекообразной. Африканский ископаемый род Proconsul, например, тоже вполне может считаться человекообразной, а не обычной, обезьяной, потому что после разделения двух ветвей в момент рандеву № 5 он оказался на стороне человекообразных обезьян. Но то, что он относится к линии человекообразных обезьян, еще не проясняет вопрос о наличии хвоста. (Данные в совокупности указывают на то, что у проконсула хвоста не было.)
Как же называть животных, которые жили между сопредком № 5 и проконсулом и еще не утратили хвост? Строгий последователь кладистического подхода назвал бы их человекообразными обезьянами, потому что они принадлежат к их ветви. Какой-нибудь другой специалист по систематике назвал бы их обычными обезьянами, потому что у них есть хвост. А я повторю: глупо зацикливаться на названиях.
Обезьяны Старого Света, Cercopithecidae, образуют кладу, то есть группу, которая включает всех потомков одного предка. А вот обезьяны как таковые (monkeys), к которым относятся обезьяны Старого Света и обезьяны Нового Света, кладу не образуют, потому что происходят от разных предков. Обезьяны Старого Света ближе к человекообразным обезьянам, чем к обезьянам Нового Света. Вместе с человекообразными обезьяны Старого Света образуют группу узконосые обезьяны (Catarrhini). И, наконец, все три группы – обезьяны Старого Света, обезьяны Нового Света и человекообразные – составляют кладу Anthropoidea. Обезьяны Старого и Нового Света (monkeys) образуют искусственную (парафилетическую) группу, потому что она включает всех широконосых и отчасти узконосые обезьяны – за исключением человекообразных. Вообще обезьян Старого Света было бы удобнее называть хвостатыми человекообразными обезьянами. Как я уже говорил, Catarrhme означает “направленный вниз нос”. В этом отношении мы – достойные представители Catarrhme. Вольтеровский доктор Панглосс как-то заметил, что “носы созданы для очков, вот мы и стали носить очки”. Он мог бы добавить, что наши направленные вниз ноздри препятствуют попаданию в нос дождя. А Platyrrhme означает “плоский (широкий) нос”. Таким образом, носы стали не только отличительным признаком двух крупнейших групп приматов, но и дали им названия.
Рандеву № 6
Обезьяны Нового Света
Рандеву № 6 назначено около 40 млн лет назад. Здесь широконосые обезьяны Нового Света (Platyrrhini) встречаются с нами, людьми, и с первым антропоидом – сопредком № 6, нашим прародителем примерно в трехмиллионном поколении. Землю покрывали густые тропические леса, и даже Антарктида (по крайней мере частично) была зеленой. Хотя сейчас все широконосые обезьяны живут в Южной или Центральной Америке, рандеву № 6 почти наверняка произошло не там. Полагаю, это случилось в Африке. Группа приматов с плоским носом, не оставивших в Африке ныне живущих потомков, перебралась в Южную Америку от 25 млн лет (к этому времени относятся первые ископаемые южноамериканские обезьяны) до 40 млн лет назад (рандеву № 6). В то время Южная Америка и Африка находились друг к другу ближе, чем теперь, а уровень моря был ниже. Поэтому между Западной Африкой и Южной Америкой могла существовать цепь островов, и обезьяны переправлялись с одного острова на другой на плотах из фрагментов мангровых растений – на них вполне можно жить некоторое время. Направления течений в те времена были подходящими для таких спонтанных путешествий. Другая крупная группа животных, дикобразоподобные грызуны ((Hystncognatha), судя по всему, прибыла в Южную Америку примерно тогда же. Причем, похоже, они тоже эмигрировали из Африки: их даже назвали в честь африканского дикобраза – Hystnx. Возможно, обезьяны переправились в Африку по той же цепи островов, что и грызуны, и пользовались теми же течениями. Хотя они путешествовали, вероятно, на разных плавсредствах.
Значит ли это, что приматы Нового Света произошли от одной-единственной предковой популяции, эмигрировавшей из Африки?
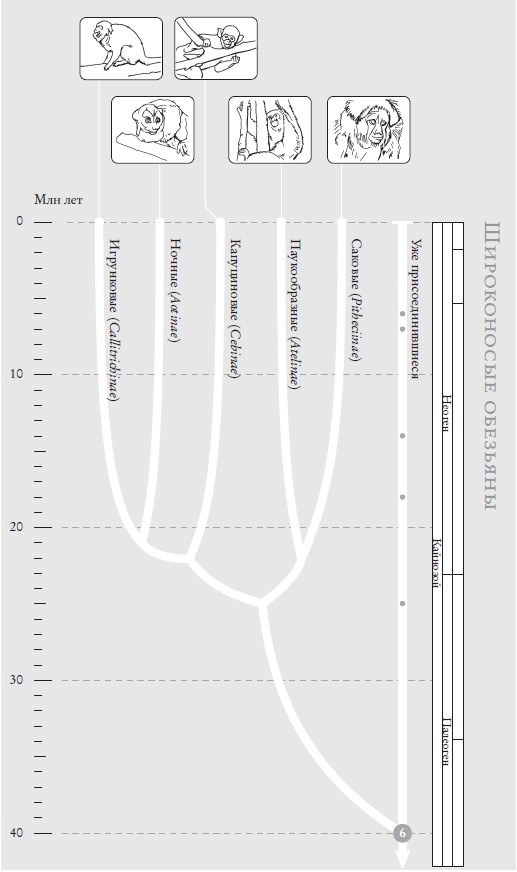
Широконосые обезьяны. Филогенетическое древо примерно для ста видов обезьян Нового Света. Филогения обезьян до сих пор вызывает разногласия, и я привожу общепринятую схему.
На рис. (слева направо): золотистая игрунка (Leontopithecus rosalia); мирикина (Aotus trivirgatus); беличий саймири, или обыкновенная беличья обезьяна (Saimiri sciureus); черный ревун (Alouatta caraya); саки-монах (Pithecia monachus).
Или приматы переправлялись с материка на материк неоднократно? А если так, можно ли это доказать? Если говорить о грызунах, то в Африке до сих пор живут дикобразоподобные грызуны – в том числе африканские дикобразы, землекоповые, скальные крысы и камышовые хомячки. Если бы оказалось, что некоторые южноамериканские грызуны – близкие родственники одной африканской группы (например дикобразов), а другие – близкие родственники другой (например землекоповых), это явилось бы доказательством того, что грызуны эмигрировали в Южную Америку не однажды. Однако не похоже, что южноамериканские грызуны происходят от разных предков, и, скорее всего, миграция грызунов стала однократным событием. Впрочем, это доказательство не то чтобы очень убедительно. Что касается приматов Южной Америки, то они тоже гораздо ближе друг к другу, чем к любому африканскому примату. Это говорит и о том, что с высокой вероятностью волна миграции приматов была единичной. Но, опять-таки, это не самое убедительное доказательство.
Пользуясь случаем, напомню, что неправдоподобие истории о переправе на плотах – еще не повод сомневаться в ней. Странно, правда? Ведь в обыденной жизни все наоборот: чем невероятнее история, тем больше у нас оснований полагать, что она не могла произойти. Однако с вопросом межконтинентальной переправы обезьян, грызунов или кого угодно дело обстоит немного иначе. Ведь переправа должна была произойти лишь однажды, а времени на это было гораздо больше, чем мы можем представить. Вероятность того, что плот с беременной самкой обезьяны достиг противоположного берега, составляет 1: 10000. В рамках человеческой жизни такая вероятность равна нулю. Однако если в нашем распоряжении 10 млн лет, это событие становится почти неизбежным. Оно должно было произойти всего один раз, а после этого все было просто. Счастливая самка родила детей, и они основали династию, которая в итоге дала все разнообразие обезьян Нового Света. Пословица гласит, что большие дубы вырастают из маленьких желудей.
Как бы то ни было, случайные переправы на плотах не так уж редки. Мелких животных довольно часто можно увидеть дрейфующими по морю. А иногда – не очень маленьких. Длина зеленой игуаны может достигать метра, а то и двух. Процитирую заметку Элен Ценски и ее соавторов, опубликованную в журнале “Нейчур”:
Четвертого октября 1995 года не менее пятнадцати особей зеленой игуаны (Iguana iguana) появилось на восточном побережье карибского острова Ангилья. Ранее этот вид на острове не встречался. Игуаны прибыли на плоту из вырванных с корнем деревьев (длина некоторых превышала девять метров). Местные рыбаки говорят, что плот был очень велик, и им потребовалось два дня, чтобы разобрать его. Они рассказали, что видели игуан на берегу и на плавающих стволах в бухте.
По-видимому, игуаны, жившие на другом острове, устроились на ночлег в деревьях, которые унес в море ураган – либо “Луис”, пронесшийся над Восточными Карибами 4–5 сентября, либо “Мэрилин”, который бушевал две недели спустя. Ни один из этих ураганов не прошел над Ангильей. Впоследствии Ценски и ее коллеги ловили или наблюдали зеленых игуан на Ангилье и на островке в полукилометре от берега. В 1998 году популяция зеленых игуан, включавшая по меньшей мере одну фертильную самку, еще обитала на Ангилье. Заметим, что игуаны и родственные им ящерицы отлично колонизируют острова. Игуаны обитают даже на Фиджи и Тонга – а ведь эти места гораздо труднодоступнее, чем острова Вест-Индии.
Кроме передвижения по ветвям на четырех конечностях, как делают многие обезьяны Старого Света, некоторые обезьяны Нового Света умеют висеть на ветках, как гиббоны, и даже пользоваться брахиацией. У всех обезьян Нового Света длинный хвост, а у паукообразных обезьян, шерстистых обезьян и ревунов он очень цепкий и служит дополнительной конечностью. Эти обезьяны могут спокойно висеть на одном хвосте – или на руке, ноге и хвосте (в любом сочетании). Наблюдая за паукообразной обезьяной, так и ждешь, что на конце ее хвоста найдутся несколько цепких пальцев[17].
Среди обезьян Нового Света есть виртуозные прыгуны. Кроме того, здесь обитают совиные обезьяны – единственная группа антропоидов, ведущая ночной образ жизни (у них большие, как у кошек или сов, глаза – таких нет больше ни у кого из обезьян Старого или Нового Света), и карликовые игрунки размером с соню – самые мелкие из антропоидов. Однако самые крупные из обезьян Нового Света – обезьяны-ревуны – по размеру напоминают крупных гиббонов. Ревуны похожи на гиббонов способностью раскачиваться на руках. Кроме того, и ревуны, и гиббоны – очень шумные животные. Однако если крики гиббонов напоминают сирены полицейских машин, то группа ревунов с их раздутыми горловыми мешками, которые служат резонаторами, скорее напоминает эскадрилью реактивных истребителей, с ревом несущихся над деревьями.
Рассказ Ревуна[18]
Новые гены появляются не из воздуха. Они образуются при дупликации прежних генов. После этого каждый из них идет своим путем, приобретая мутации и подвергаясь действию естественного отбора и дрейфа генов. Обычно мы не видим, как это происходит, но, подобно следователям на месте преступления, можем попытаться воссоздать произошедшее на основе улик. Замечательный пример – гены, ответственные за цветовое зрение.
Млекопитающие очень долго были ночными существами. День принадлежал динозаврам, у которых, если судить по ныне живущим родственникам, было отличное цветовое зрение. Также не без оснований можно предположить, что цветовое зрение имелось и у далеких предков млекопитающих – звероподобных рептилий (тероморфов), населявших Землю до динозавров. Однако за время, которое млекопитающие провели во тьме, их глаза должны были научиться улавливать каждый доступный фотон. Неудивительно, что способность различать цвета они почти утратили (о причинах мы поговорим в “Рассказе Слепой пещерной рыбы”). До сих пор у большинства млекопитающих (даже тех, которые вернулись к дневному образу жизни) слабое цветовое зрение – двухроматическое. Речь о количестве типов светочувствительных клеток – колбочек – в сетчатке. У человека и остальных узконосых обезьян, а также у обезьян Старого Света, три типа колбочек: красные, зеленые и синие. То есть у нас зрение трихроматическое. Однако есть данные, что мы заново приобрели третий тип колбочек после того, как его утратили наши ночные предки. У большинства других позвоночных, например рыб или рептилий (но не млекопитающих!), зрение трихроматическое (имеются колбочки трех типов) или тетрахроматическое (четыре типа). У птиц и черепах зрение бывает еще сложнее. Что касается обезьян Нового Света, особенно ревунов, то они в особенном положении.
Есть указания на то, что австралийские сумчатые, в отличие от большинства млекопитающих, обладают хорошим трихроматическим зрением. Кэтрин Аррес и ее коллеги, открывшие его у поссумов-медоедов и австралийских сумчатых землероек (такое зрение было найдено и у кенгуру-валлаби), предположили, что австралийские (но не американские) сумчатые сохранили древний светочувствительный пигмент рептилий, утраченный остальными млекопитающими. Однако в целом у млекопитающих все-таки самое слабое среди позвоночных цветовое зрение. Большинство млекопитающих если и различает цвета, то примерно на уровне страдающих дальтонизмом людей. Любопытные исключения из этого правила можно найти среди приматов. Неслучайно они чаще других млекопитающих используют для брачной демонстрации яркие цвета.
Австралийские сумчатые, в отличие от нас, видимо, никогда не теряли цветовое зрение. Мы же, судя по своим родственникам среди млекопитающих, скорее всего, не унаследовали от рептилий трихроматическое зрение, а заново изобрели его. Причем два раза: первый раз это сделали обезьяны Старого Света и человекообразные обезьяны, а второй – ревуны (только они, а не все обезьяны Нового Света). Цветовое зрение ревунов похоже на цветовое зрение человекообразных обезьян, однако отличается от него достаточно, чтобы понять: возникло оно независимо.
Что же такого хорошего в цветовом зрении, что трихроматизм возник независимо у обезьян Старого и Нового Света? Основная гипотеза гласит, что цветовое зрение связано с питанием фруктами. На фоне однотонной зеленой листвы фрукты выделяются. Это, в свою очередь, не случайно. Фрукты скорее всего приобрели яркие цвета для привлечения поедающих плоды животных, например обезьян, которые играют важную роль в распространении семян. Кроме того, трихроматическое зрение помогает находить на фоне темной листвы более молодые и мясистые листья (которые бывают бледно-зелеными, иногда даже красными). Впрочем, последнее вряд ли идет на пользу растениям.
Цвет завораживает нас. Слова, обозначающие цвета – первые прилагательные, которые узнают дети, и именно эти слова они раньше всего пытаются связать с каким-нибудь подходящим существительным. Но мы редко вспоминаем, что цвета и оттенки, которые мы видим – это ярлыки, которые наше сознание навешивает на участки спектра, слегка различающиеся длиной электромагнитных волн. Красный свет соответствует длине волны около 700 нм, фиолетовый – около 420 нм. При этом видимое излучение в этих рамках составляет ничтожную часть спектра. Ведь длина волны может варьировать от нескольких километров (некоторые радиоволны) до долей нанометра (гамма-излучение).
Все глаза на планете устроены так, чтобы воспринимать волны тех длин, в которых наша звезда ярче всего и которые проходят сквозь земную атмосферу. Однако способность глаза к восприятию электромагнитных волн ограничена биохимическими системами, необходимыми для улавливания размытого диапазона излучения. Законы физики устанавливают для видимого диапазона длин волн более четкие границы. Ни одно животное не воспринимает всю инфракрасную область спектра. Ближе всего подошли к этому гремучие змеи. У них на голове есть ямки, которые хотя и не могут создавать сфокусированное изображение в инфракрасном диапазоне, дают змеям некоторую направленную чувствительность к теплу, исходящему от добычи. Также ни одно животное не может воспринимать весь диапазон ультрафиолетовой части спектра, хотя некоторые животные, например пчелы, видят в ультрафиолете немного лучше нас. Зато пчелы не видят “нашего” красного цвета: для них он инфракрасный. В целом для животных “свет” – это узкий спектр электромагнитных волн между короткими волнами ультрафиолетового излучения и длинными – инфракрасного. Различие между пчелами, людьми и змеями лишь в том, где проходят границы воспринимаемого спектра.
Более узки эти границы для каждого типа светочувствительных клеток сетчатки. Одни колбочки немного лучше воспринимают красный участок спектра, другие – синий. Цветовое зрение представляет собой сопоставление данных, полученных от различных колбочек, и качество такого зрения сильно зависит от того, сколько типов колбочек в сетчатке. В сетчатке животных с дихроматическим зрением – два типа колбочек, трихроматическим зрением – три, тетрахроматическим зрением – четыре. Каждая колбочка характеризуется кривой чувствительности, у которой есть пик и не очень симметричные “угасающие” концы. За пределами, очерченными этой кривой, клетка “слепа”.
Допустим, колбочка имеет пик в зеленой части спектра. Значит ли это, что такая клетка посылает сигналы в мозг лишь тогда, когда она “смотрит” на зеленый объект, например на траву или бильярдный стол? Безусловно, нет. Просто клетке необходимо больше красного света (в этом случае), чтобы выдавать импульсы той же интенсивности, как при заданном количестве зеленого света. Такая клетка будет одинаково реагировать как на яркий красный свет, так и на тусклый зеленый[19]. Нервная система может определить цвет объекта лишь путем сопоставления одновременно поступающих импульсов по меньшей мере от двух клеток, чувствительных к разным участкам спектра. Каждая клетка при этом “контролирует” другую. Если же клеток не две, а три, представление о цвете окажется еще полнее.
Цветное телевидение и компьютерные мониторы работают в трехцветной системе, потому что были разработаны для трихроматических глаз. В обычном мониторе каждый пиксель состоит из трех точек, расположенных настолько близко друг к другу, что глаз не может их различить. Каждая точка всегда светится одним цветом. Посмотрев на экран под достаточно большим увеличением, мы увидим одни и те же три цвета – обычно красный, зеленый и синий, хотя иногда могут использоваться и другие комбинации. Любой оттенок можно получить, регулируя интенсивность свечения трех основных цветов. Должно быть, черепах с их тетрахроматическим зрением наши телевизоры разочаровывают.
Сопоставляя интенсивность импульсов всего от трех типов колбочек, наш мозг может воспринимать огромный диапазон оттенков. Но большинство плацентарных млекопитающих обладает дихроматическим зрением: в их сетчатке колбочки двух типов. У колбочек одного типа пик восприимчивости в фиолетовой области спектра (в некоторых случаях – в ультрафиолетовой), у второго типа – где-то между зеленой и красной областями. У нас, животных с трихроматическим зрением, пик “коротковолновых” колбочек находится между фиолетовой и синей областями спектра. Такие колбочки называют синими… Другие два типа наших колбочек – так называемые зеленые и красные. Правда, даже у красных колбочек пик приходится скорее на желтоватую, чем на красную часть спектра. Но в целом кривая их чувствительности сдвинута в красную часть спектра. Так что, несмотря на то, что пик в желтой области, эти колбочки все равно генерируют сильный импульс в ответ на красный свет. Поэтому, если вычесть интенсивность импульса от зеленых колбочек из интенсивности импульса красных колбочек, мы получим особенно сильный сигнал в ответ на красный свет.
Кроме колбочек, в сетчатке еще есть палочки – светочувствительные клетки, которые отличаются от колбочек формой и особенно эффективны ночью. В цветовом зрении эти клетки не участвуют, и мы больше не будем о них говорить.
Химия и генетика цветового зрения довольно хорошо изучены. Главными молекулярными акторами здесь выступают опсины. Это молекулы белка, которые в колбочках и палочках выполняют функцию зрительных пигментов. Каждая молекула опсина связана с одной молекулой ретиналя – химического соединения, которое является производным витамина А [20]. Молекула ретиналя скручена в петлю, которая встраивается в молекулу опсина. При попадании фотона с подходящей длиной волны узел распрямляется. Это служит сигналом для клетки: она посылает нервный импульс, который говорит мозгу: “Вижу свет своего типа”. Тогда молекула опсина связывается с новой скрученной молекулой ретиналя, поступающей из внутриклеточных запасов.
Не все молекулы опсина одинаковы. Опсины, как и все белки, кодируются генами. Различия в ДНК приводят к образованию опсинов, чувствительных к разным цветам, и это служит генетической основой дихроматического и трихроматического зрения. А поскольку каждая клетка организма имеет полный набор генов, различия между красными и синими колбочками не в том, какие гены у них есть, а в том, какие гены работают. На этот счет есть правило: каждая колбочка включает лишь один тип генов.
Гены, кодирующие наши зеленый и красный опсины, очень похожи друг на друга и находятся на Х-хромосоме (половая хромосома, которая у женщин есть в двух копиях, у мужчин – в одной). Ген, отвечающий за образование синего опсина, немного от них отличается и находится не на половой хромосоме, а на одной из обычных хромосом, которые называют аутосомами (в нашем случае это седьмая хромосома). Наши зеленые и красные клетки явно образовались в результате недавней дупликации гена, который, в свою очередь, задолго до этого образовался в результате дупликации гена синего опсина. Тип зрения животного – дихроматический или трихроматический – зависит от того, сколько разных генов опсинов у него в геноме. Если в геноме животного есть, например, гены синего и зеленого опсинов, но отсутствует ген красного опсина, такое животное будет дихроматом.
Прежде чем заняться ревунами и историей приобретения ими трихроматического зрения, попытаемся понять, как устроено странное для нас дихроматическое зрение остальных обезьян Нового Света. Кстати, дихроматическое зрение есть у некоторых лемуров, но не у всех обезьян Нового Света (у ночных обезьян зрение монохроматическое). Так что пока забудем о ревунах и других необычных видах.
Во-первых, исключим из обсуждения “синий” ген, который находится на аутосомах всех особей независимо от пола. С “красными” и “зелеными” генами на Х-хромосоме все сложнее и гораздо интереснее. На каждой Х-хромосоме есть только один локус, в котором может находиться красный или зеленый аллель[21]. Поскольку у самки две Х-хромосомы, она может обладать и красным, и зеленым аллелями одновременно. Однако у самца с его единственной Х-хромосомой может быть лишь один аллель: красный либо зеленый. Таким образом, типичный самец обезьяны Нового Света должен обладать дихроматическим зрением, потому что у него колбочки лишь двух типов: синие плюс красные либо зеленые. По нашим меркам, все эти самцы – дальтоники. Однако это дальтоники двух типов: у одних самцов в популяции нет зеленого опсина, у других – красного. Синий есть у всех.




