



Ричард Докинз
Рассказ предка. Паломничество к истокам жизни
Рассказ Бонобо
Бонобо (Pan paniscus) очень похож на обыкновенного шимпанзе (Pan troglodytes), и до 1929 года их считали единым видом. По размеру бонобо, несмотря на свое второе название, от которого давно следует отказаться, – карликовый шимпанзе, – не меньше обычного. У него немного другие пропорции тела и особенности поведения, и это послужит темой его рассказа. Приматолог Франс де Вааль выразился так: “Шимпанзе решает проблемы секса, используя власть; бонобо решает проблемы власти, используя секс”. Бонобо действительно пользуются сексом как валютой социального взаимодействия – так же, как мы пользуемся деньгами. Они используют копуляцию или имитирующие половой акт движения для примирения, для демонстрации доминирования и укрепления социальных связей с особями любого возраста и пола, включая детенышей. Педофилия бонобо совершенно не смущает. Им вообще нравятся всевозможные – филии. Де Вааль описывает, как в группе бонобо, за которыми он наблюдал в неволе, у самцов возникала эрекция, когда служащий подходил к ним во время кормления. По мнению де Вааля, так они готовятся к дележу пищи: инструментом такого дележа опять-таки является секс. Самки бонобо практикуют GG-трение (трение гениталиями):
Одна самка цепляется руками и ногами за партнершу, которая, стоя на руках и ногах, приподнимает ее с земли. Затем они трутся гениталиями, издавая при этом смешки и визги, которые, вероятно, выражают оргазменные ощущения.
Изображение любвеобильных бонобо а-ля Хейт-Эшбери привело к неразберихе в головах вполне порядочных людей, взросление которых пришлось на 60-е годы. Или, возможно, эти люди просто приняли близко к сердцу средневековые бестиарии, согласно которым животные существуют лишь для того, чтобы преподать нам некий моральный урок. Ошибка заключается в ложном представлении о том, что мы ближе к бонобо, чем к обыкновенному шимпанзе. Соблазненные Маргарет Мид, мы почему-то считаем, что эта приятная и легкомысленная модель поведения нам ближе, чем патриархальный и кровожадный общественный строй обыкновенных шимпанзе. Должен вас огорчить: у нас одинаковая степень родства с обоими видами – просто потому, что Pan troglodytes и P paniscus происходят от общего предка, который жил позднее, чем наш с ними общий предок. Молекулярные данные также говорят, что шимпанзе и бонобо ближе к людям, чем к гориллам. Следовательно, люди близки к гориллам настолько же, насколько они близки к шимпанзе и бонобо. То же с орангутанами: с ними у нас та же степень родства, как и с гориллой, обыкновенным шимпанзе и бонобо.
Впрочем, отсюда не следует, что внешне мы в равной степени похожи на обыкновенного шимпанзе и бонобо. Шимпанзе со времени жизни сопредка № 1 изменились сильнее, чем бонобо, а мы скорее похожи на бонобо, чем на шимпанзе, и наоборот. При желании можно найти общие черты с обоими нашими родственниками из рода Pan – возможно, их примерно поровну. С обоими этими видами нас связывает одинаковая степень родства, потому что у нас общий предок. В этом и заключается мораль “Рассказа Бонобо” – и не только его.
Рандеву № 2
Гориллы
“Молекулярные часы” показывают, что рандеву № 2, где к нам присоединяются гориллы, снова назначено в Африке. От рандеву № 1 его отделяет миллион лет. Семь миллионов лет назад Северная и Южная Америка еще не срослись, Гималаи только что поднялись, а Анды – еще нет. При этом континенты выглядели почти так, как сейчас. А климат Африки, хотя и был чуть более влажным и имел менее выраженную сезонность, в целом был похож на нынешний. В Африке было гораздо больше лесов. Даже Сахара была лесистой саванной.
К сожалению, не осталось ископаемых, которые могли бы заполнить пробел между сопредками №№ 1 и 2, и мы не знаем, был ли сопредок № 2 (наш предок примерно в 300-тысячном поколении) похож на гориллу, на шимпанзе или на человека. Мне кажется, он был похож на шимпанзе – просто потому, что гориллы выглядят очень своеобразно. Впрочем, не стоит преувеличивать своеобразие горилл. Это не самые крупные обезьяны в истории Земли. Похожая на орангутана азиатская человекообразная обезьяна гигантопитек (Gigantopithecus) была на две головы выше самой крупной из горилл и гораздо шире ее в плечах. Гигантопитек жил на территории нынешнего Китая и вымер всего 500 тыс. лет назад. Таким образом, он отчасти современник Homo erectus и архаичного Homo sapiens. Это было недавно, и некоторые фантазеры предположили, что йети… Нет-нет, я отказываюсь это обсуждать. Гигантопитек, по-видимому, передвигался на четырех конечностях, как и гориллы и шимпанзе – но не как орангутаны: последние живут в основном на деревьях.
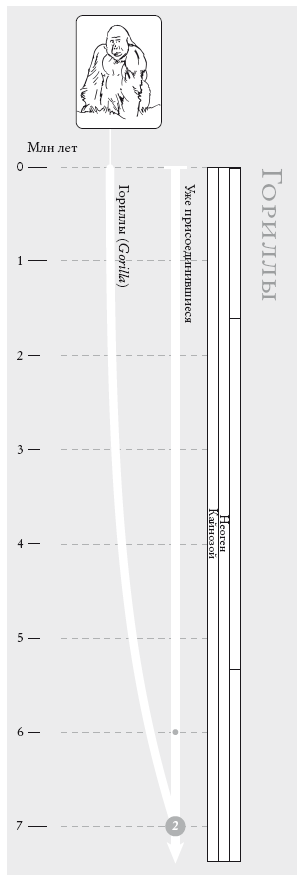
Гориллы. Филогенетическое древо, иллюстрирующее отход горилл от других африканских человекообразных обезьян. Согласно генетическим данным, это произошло ок. 7 млн лет назад. Правая ветвь – шимпанзе и люди (сопредок № 1 отмечен точкой, соответствующей 6 млн лет), левая – единственный вид горилл (который, как теперь считается, следует делить на два). На рис.: западная горилла (Gorilla gorilla).
Логично предположить, что сопредок № 2, как и шимпанзе, ходил, опираясь на костяшки пальцев, и на ночь забирался на дерево. Естественный отбор в условиях тропического солнца благоприятствует сильной пигментации, которая защищает от ультрафиолетового излучения. Так что сопредок № 2, скорее всего, был темно-коричневым или черным. А поскольку все обезьяны, кроме людей, покрыты волосами, было бы странно, если бы сопредки №№ 1 и 2 сильно от них отличались. И, наконец, поскольку шимпанзе, бонобо и гориллы живут в лесу, рандеву № 2 разумно назначить там же. Лес этот должен находиться в Африке, но где именно, к сожалению, неясно.
Гориллы отличаются от шимпанзе не только размерами. Это нужно иметь в виду при реконструкции сопредка № 2. Гориллы – строгие вегетарианцы. У самцов есть гаремы. Шимпанзе беспорядочны в половых связях, и эти различия в системах скрещивания любопытным образом сказываются на размере их яичек. (Об этом мы узнаем из “Рассказа Тюленя”.) Я подозреваю, что системы скрещивания вообще довольно лабильны, то есть в процессе эволюции легко изменяются. Пока я не вижу способа узнать, как с этим обстояло дело у сопредка № 2. Ведь, например, у людей в разных культурах разные системы скрещивания – от строгой моногамии до огромных гаремов. Поэтому я не вижу смысла гадать, как размножался сопредок № 2, и закрываю эту тему.
Рассказ Гориллы
Становление дарвинизма, которое пришлось на XIX век, привело к появлению двух противоположных взглядов на человекообразных обезьян. Противники Дарвина, хотя и согласившиеся принять идею эволюции, были в ужасе от возможного родства с грубыми, отвратительными тварями и отчаянно пытались преувеличить наши отличия. Особенно это касалось горилл. Человекообразных обезьян называли “животными”, противопоставляя их человеку. Более того: если, например, кошки и олени кажутся красивыми, гориллы и другие человекообразные обезьяны именно в силу сходства с нами кажутся карикатурами, пародиями.
Дарвин часто напоминал, что он думает по этому поводу: так, в “Происхождении человека” он как бы между прочим отмечает, что обезьяны “с удовольствием курят табак”. Томас Генри Гексли (Хаксли), знаменитый последователь Дарвина, однажды участвовал в ожесточенном споре с сэром Ричардом Оуэном, ведущим анатомом того времени. Оуэн утверждал (ошибочно, как доказал Гексли), что наличие малого гиппокампа является уникальной чертой человеческого мозга. Однако сейчас ученым уже не кажется, что мы напоминаем человекообразных обезьян. Они знают, что мы относимся к африканским человекообразным обезьянам. И теперь мы противопоставляем человекообразных обезьян (включая человека) всем остальным обезьянам.
Так было не всегда. Прежде человекообразных обезьян объединяли с другими высшими приматами, и в некоторых ранних описаниях к человекообразным обезьянам относят, например, павианов или маготов. Удивительно другое. Задолго до того, как люди стали мыслить в терминах эволюции, и до того, как человекообразных обезьян противопоставили остальным обезьянам, крупных человекообразных обезьян нередко путали с людьми. И хотя приятно думать, что такое представление предвосхищало идею эволюции, следует признать, что оно сродни расизму. Первые европейские путешественники видели в шимпанзе и гориллах Африки близких родственников чернокожих – но не белых. Интересно, что у племен Юго-Восточной Азии и Африки есть легенды, в которых эволюция как бы направлена вспять: согласно преданиям, крупные человекообразные обезьяны – это согрешившие люди. “Орангутан” по-малайски значит “лесной человек”.
Рисунок орангутана (Ourang Outang), выполненный в 1658 году голландским доктором Бонциусом, по словам Гексли, изображает “не что иное, как очень волосатую женщину, довольно миловидную, с совершенно человеческими пропорциями тела и ногами”. Правда, почему-то волосы у нее растут по всему телу, кроме одного из участков, которые у женщин как раз покрыты волосами: лобок, как видно на рисунке, гол. Весьма антропоморфны и рисунки, сделанные Хоппиусом (1763), учеником Линнея. У одного из изображенных существ есть хвост, однако в остальном оно неотличимо от человека: стоит на двух ногах и держит трость. Плиний Старший писал, что “хвостатых существ, говорят, даже видели за игрой в шашки”.
Хочется думать, что все эти мифы должны были подготовить человечество к идее эволюции или даже ускорить ее открытие в XIX веке. Однако это не так. Вместо этого возникла путаница между человекообразными обезьянами, обычными обезьянами и человеком. Из-за этого бывает трудно понять, когда был открыт тот или иной вид человекообразных обезьян – а часто невозможно понять даже, о каком виде речь. Исключением является горилла, которая стала известна науке в последнюю очередь.
В 1847 году американский миссионер доктор Томас Сэвидж увидел в доме другого миссионера на реке Габон “череп, по словам туземцев, принадлежащий существу, напоминающему обезьяну и замечательному своими размерами, свирепостью и повадками”. В том же году, в котором вышло “Происхождение видов”, газета “Иллюстрей-тед Лондон ньюс” напечатала сенсационную статью о гориллах. Эти вымыслы (в том числе об их свирепости, достигшие максимального масштаба в фильмах о Кинг-Конге) не сравнятся даже с россказнями путешественников того времени:
Наблюдать горилл вблизи почти невозможно – особенно потому, что, увидев человека, они сразу нападают. Взрослый самец обладает невероятной силой, зубы у него крупные и мощные. Говорят, что горилла, спрятавшись в толстых ветвях деревьев, подкарауливает человека. Когда он проходит под деревом, горилла спускает страшную заднюю ногу… и хватает жертву за горло, поднимая в воздух, после чего швыряет на землю мертвой. Этим животным движет чистая злоба: оно не ест человеческую плоть, а просто получает жестокое удовольствие от убийства.
Сэвидж решил, что череп, который он видел у миссионера, принадлежит “новому виду орангутана”. Позднее он пришел к выводу, что новый вид – не что иное, как “понго” из рассказов первых путешественников по Африке. Давая виду название, Сэвидж с коллегой-анатомом, профессором Уайменом, решил воздержаться от “понго” и выбрал давно забытое слово “горилла”: так карфагенский мореплаватель назвал диких волосатых людей, якобы встреченных им на острове у побережья Африки. И словом “горилла” стали обозначать найденное Сэвиджем животное – на латыни и других языках. А “понго” впоследствии стало латинским родовым названием азиатских орангутанов.
Судя по тому, где Сэвидж обнаружил животное, это скорее всего была западная горилла (Gorilla gorilla). Сэвидж и Уаймен отнесли его к тому же роду, что и шимпанзе, дав название Troglodytes gorilla. Однако по правилам зоологической номенклатуры на имя Troglodytes не имели права ни шимпанзе, ни горилла, поскольку оно уже принадлежало – ни за что не угадаете! – крошечной птице крапивнику. Это слово сохранилось в качестве видового названия обыкновенного шимпанзе – Pan troglodytes. А видовое название, которое дал горилле Сэвидж, возвели до родового названия Gorilla. Так называемая горная горилла в 1902 году была “открыта” – точнее, застрелена – немцем Робертом фон Берингом. Теперь ее считают подвидом восточной гориллы, а сама восточная горилла названа в его честь: Gorilla beringei. Не уверен, что это справедливо.
Сэвидж не верил, что его горилла и правда принадлежит к расе, о которой упоминал карфагенянин. Однако есть и другой пример: у Геродота и Гомера упоминаются “пигмеи” – легендарная раса крошечных людей. В XVII и XVIII веках ученые пришли к выводу, что это шимпанзе, к тому времени открытые в Африке. У Тайсона (1699) есть рисунок “пигмея”, который, как обратил внимание Гексли, изображает молодого шимпанзе, пусть и поставленного на две ноги и опирающегося на трость. Ну а сегодня мы снова используем слово “пигмей”" для обозначения очень маленького человека.
Это возвращает нас к мысли о расизме, которым до недавнего времени отличалась наша культура. Ранние исследователи приписывали лесным племенам более близкое родство с шимпанзе, гориллами и орангутанами, чем с собой. В XIX веке, уже после Дарвина, эволюционисты зачастую рассматривали африканцев как промежуточное звено между обезьянами и европейцами. Это неверно не только фактически, но и нарушает фундаментальный принцип эволюции. Два двоюродных брата всегда в одинаковом родстве с любым членом “внешней группы” (человеком, не принадлежащим к их группе): у них общий предок. Все люди в равном родстве со всеми гориллами (см. “Рассказ Бонобо”). Однако никуда не девается расизм и специесизм (видовой шовинизм), а также вечные сомнения в том, насколько должны быть широки рамки морали. Все это неловким и подчас жестоким образом влияет на отношение человека к нашим собратьям – людям и человекообразным обезьянам[10].
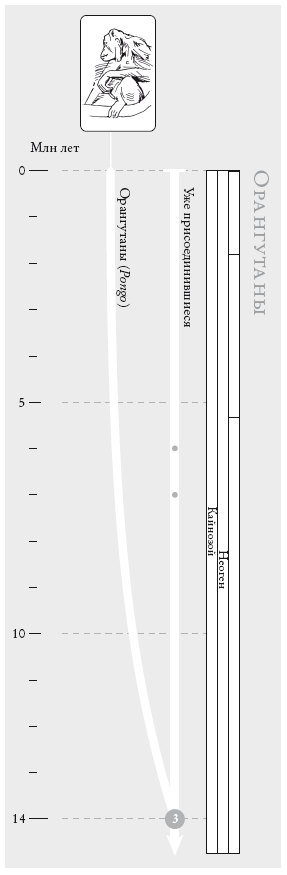
Орангутаны. Согласно общепринятой точке зрения, два вида азиатских орангутанов отделились от остальных человекообразных обезьян ок. 14 млн лет назад. Как и на остальных филогенетических схемах, правая ветвь объединяет уже присоединившихся к нам "пилигримов”. Предыдущие сопредки обозначены точками.
На рис.: борнейский орангутан (Pongo pygmaeus).
Рандеву № 3
Орангутаны
Согласно молекулярным данным, рандеву № 3 (здесь к нам присоединяется орангутан) назначено на 14 млн лет назад – на середину миоценовой эпохи. Хотя тогда на нашей планете уже начало холодать, климат все же был теплее нынешнего, а уровень моря – выше. Из-за этого (а также небольших различий в положении континентов) перешеек между Азией и Африкой, а также значительная часть Юго-Восточной Европы время от времени затапливались. Это важно в контексте размышлений о том, где мог жить сопредок № 3 – наш гипотетический предок в 666666-тысячном поколении. Жил ли он в Африке, как сопредки №№ 1 и 2 – или в Азии? Поскольку он является общим предком человека и азиатских человекообразных обезьян, следует быть готовым к встрече с ним на любом из двух континентов, и тому есть доказательства. В пользу Азии говорит множество окаменелостей, датируемых как раз второй половиной миоцена. С другой стороны, похоже, что именно в Африке появились человекообразные обезьяны – еще до миоценовой эпохи. Различные формы человекообразных обезьян процветали в Африке в раннем миоцене – проконсулиды (несколько видов человекообразных обезьян рода Proconsul) и другие, например афропитек (Afropithecus) и кениапитек (Kenyapithecus). Ближайшие современные формы, а также все наши послемиоценовые ископаемые родственники найдены к Африке.
О наших особых отношениях с шимпанзе и гориллами известно лишь несколько десятилетий. Прежде большинство антропологов считало, что человек – это сестринская группа по отношению ко всем человекообразным обезьянам и что он, таким образом, одинаково близок к африканским и к азиатским обезьянам. С общего согласия предпочтение отдали Азии, поскольку именно там жили наши последние миоценовые предки. Некоторые авторитеты даже объявили, что нашли нашего ископаемого “предка” – рамапитека (Ramapithecus). Сегодня считается, что рамапитек – то же животное, что и сивапитек (Sivapithecus), – а по законам зоологической номенклатуры приоритет имеет позднейшее название. Таким образом, название “рамапитек” больше использовать нельзя – а жаль, потому что оно прижилось. К сивапитеку (рамапитеку) ученые относятся по-разному, однако многие соглашаются в том, что он близок к линии, от которой произошел орангутан, – а может, он сам непосредственный предок орангутана. Гигантопитек (Gigantopithecus) – нечто вроде гигантского наземного сивапитека. Известны и другие азиатские ископаемые того периода. Так, за титул миоценового предка человека борются уранопитек (Ouranopithecus) и дриопитек (Dryopithecus). Если бы они жили на “правильном” континенте… Но, как мы увидим, “если” может оказаться реальностью.
Если бы позднемиоценовые человекообразные обезьяны жили в Африке, а не в Азии, в нашем распоряжении имелся бы непрерывный ряд ископаемых, связывающих современных африканских обезьян с раннемиоценовыми формами и богатой африканской фауной проконсулид. После того как с помощью молекулярных данных была абсолютно доказана близость человека к африканским шимпанзе и гориллам, а не к азиатским орангутанам, охотники за предками человека отвернулись от Азии. Ученые решили, что, как бы ни были хороши азиатские человекообразные обезьяны, наша родословная должна относиться к Африке и включать миоценовые формы, – и заключили, что по неизвестным причинам после расцвета миоценовых проконсулид наши африканские предки просто не подвергались фоссилизации.
Это положение вещей сохранялось до 1998 года, когда вышла статья Каро-Бет Стюарт и Тодда Р. Дисотелла “Эволюция приматов – в Африку и из Африки”. В статье (это замечательный пример всестороннего подхода к вопросу) рассказывается о миграциях между Африкой и Азией. И, знаете, сопредок № 3, похоже, жил в Азии. Но об этом после.
А пока задумаемся, как он выглядел. Поскольку сопредок № 3 является общим предком орангутана и современных африканских человекообразных обезьян, он должен быть похожим на любую из них – и одновременно на орангутана. Найдены ли ископаемые, которые могли бы подсказать нам ответ? Что ж, взглянем на генеалогическое древо. К нужному нам периоду относятся люфенгпитек, ореопитек, сивапитек, дриопитек и уранопитек. Наиболее вероятная реконструкция облика сопредка № 3 совместит черты всех пяти азиатских ископаемых родов (при условии, что это животное обитало в Азии). Послушаем “Рассказ Орангутана”, а там решим.
Рассказ Орангутана
Возможно, заявление о наших давних связях с Африкой было поспешным. Что если наши предки покинули Африку около 20 млн лет назад и поселились в Азии, а 10 млн лет назад вернулись в Африку?
Если так, то современные человекообразные обезьяны, включая тех, которые в итоге оказались в Африке, происходят от линии, мигрировавшей из Африки в Азию. Гиббоны и орангутаны – потомки эмигрантов, оставшихся в Азии. Поздние их потомки вернулись в Африку, где раннемиоценовые человекообразные обезьяны уже вымерли, и уже там породили горилл, шимпанзе, бонобо – и нас.
С этим вполне согласуются данные о дрейфе континентов и колебаниях уровня моря. Как раз в нужные моменты в Аравийском море имелись перешейки, соединявшие Азию с Африкой. В пользу этой теории говорит и принцип парсимонии, то есть экономии: хороша та теория, которая многое объясняет, постулируя малое. (Как я не раз говорил, по этому критерию теория естественного отбора Дарвина является, пожалуй, лучшей на свете.) В данном случае речь о том, чтобы до минимума снизить число предполагаемых миграционных событий. С этой точки зрения теория о том, что наши предки все время обитали в Африке (без миграций), на первый взгляд экономнее теории о том, что наши предки сначала переселились из Африки в Азию (первая миграция), а потом вернулись в Африку (вторая миграция).
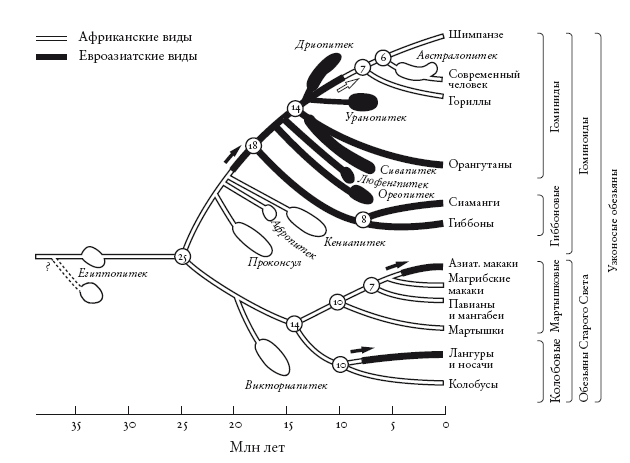
Филогенетическое древо африканских и азиатских обезьян. Расширения обозначают даты, полученные на основе анализа ископаемых, а линии, соединяющие их с древом, построены методом парсимонии. Stewart and Disotell [273].
Но это частный случай: мы рассмотрели лишь собственную линию и проигнорировали остальных человекообразных обезьян, особенно многие ископаемые виды. Стюарт и Дисотелл провели пересчет миграционных событий, в том числе необходимых для объяснения распределения человекообразных обезьян, включая вымерших. Для этого нужно сначала построить древо, отметив на нем все виды, о которых достаточно данных. Следующий шаг – указать для каждого вида, жил он в Африке или в Азии. На диаграмме Стюарта и Дисотелла азиатские ископаемые отмечены черным цветом, африканские – белым. Там представлены не все известные ископаемые, а лишь те, для которых можно определить положение. Стюарт и Дисотелл не забыли и обезьян Старого Света, которые отделились от человекообразных около 25 млн лет назад (самое наглядное различие между ними состоит в том, что первые сохранили хвосты). Миграционные события указаны стрелками.
С учетом этих ископаемых теория переселения в Азию и обратно оказывается экономнее теории о том, что наши предки все время жили в Африке. Оставим хвостатых обезьян, которые, согласно обеим теориям, участвовали в двух волнах миграции из Африки в Азию, и рассмотрим человекообразных. Теория переселения требует всего двух волн миграции:
1. Популяция человекообразных обезьян переселилась из Африки в Азию около 20 млн лет назад. От нее произошли все азиатские человекообразные обезьян, включая современных гиббонов и орангутанов.
2. Популяция человекообразных обезьян вернулась в Африку. От нее произошли современные африканские человекообразные обезьяны, включая нас.
При этом теория постоянного обитания в Африке для объяснения расселения человекообразных обезьян требует шести миграций. И все они направлены из Африки в Азию:
1. Гиббоны (около 18 млн лет назад).
2. Ореопитек (около 16 млн лет назад).
3. Люфенгпитек (около 15 млн лет назад).
4. Сивапитек и орангутан (около 14 млн лет назад).
5. Дриопитек (около 13 млн лет назад).
6. Уранопитек (около 12 млн лет назад).
Разумеется, эти подсчеты верны лишь при условии, что Огюарт и Дис-отелл, опиравшиеся на анатомические данные, верно построили древо. Они, например, считают, что ближайший из всех ископаемых родственник современных африканских человекообразных обезьян – это уранопитек. Его линия ответвляется непосредственно перед линией африканских человекообразных обезьян. Все следующие родственники, согласно оценкам Стюарт и Дисотелла, – азиаты (дриопитек, сивапитек и другие). Если же авторы неверно интерпретировали анатомические данные (например, если ближайший родственник современных африканских человекообразных обезьян – на самом деле ископаемый африканский кениапитек), подсчет миграционных событий надо провести заново.
Сама генеалогическая схема тоже построена по принципу парсимонии. Но мы не пытаемся уменьшить число миграционных событий, необходимых для теории, и игнорируем географию. Вместо этого мы хотим минимизировать число анатомических совпадений (конвергентная эволюция), положенных в основу теории. Так мы получаем древо, не учитывающее географические события, и, чтобы подсчитать миграционные события, накладываем географические данные (черная и белая маркировка на диаграмме). И приходим к выводу: с наибольшей вероятностью современные африканские человекообразные обезьяны – гориллы, шимпанзе и люди – пришли из Азии.
А теперь любопытное замечание. Ричард Клейн из Стэнфордского университета в учебнике по эволюции человека приводит обзор анатомических данных основных ископаемых. Клейн, сравнивая азиатского уранопитека и африканского кениапитека, задается вопросом, кто из них сильнее похож на нашего близкого предка (или родственника) австралопитека. И делает вывод: австралопитек сильнее похож на уранопитека, и если бы уранопитек жил в Африке, он вполне мог быть предком человека. Однако “на основе географических и морфологических данных”, пишет Клейн, кениапитек более подходящий кандидат. Видите? Клейн предполагает, что африканские человекообразные обезьяны просто не могли произойти от азиатского предка, даже если анатомические данные указывают на это. Таким образом, он подсознательно предпочитает географическую парсимонию анатомической. Из соображений анатомической парсимонии уранопитек к нам ближе, чем кениапитек. Однако, хотя об этом не говорится открыто, географическая парсимония берет верх. Стюарт и Дисотелл утверждают, что при рассмотрении географического распространения всех известных ископаемых анатомическая и географическая парсимонии согласуются. То есть географические данные не противоречат утверждению Клейна, на основе анатомических данных признавшему: уранопитек ближе к австралопитеку.
Возможно, этот спор далек от разрешения. Не так-то просто манипулировать анатомической и географической парсимониями. Статья Стюарт и Дисотелла вызвала бурную реакцию: в научных журналах появлялись и восторженные, и разгромные отклики. Мне кажется, что сегодня, оценив все доступные данные, мы должны отдать предпочтение теории “переселения в Азию и обратно”. Два миграционных события – меньше, чем шесть. Кроме того, мне кажется, между позднемиоценовыми азиатскими обезьянами и “нашей” линией африканских обезьян (например австралопитеками и шимпанзе) есть немалое сходство. Таким образом, теория представляет собой компромисс. Исходя из этого, я назначаю рандеву № 3 (и № 4) не в Африке, а в Азии.
Мораль “Рассказа Орангутана” неоднозначна. Когда ученый выбирает между двумя теориями, на первом месте всегда принцип парсимонии. Однако не всегда ясно, как ее оценивать. Наличие хорошей родословной бывает необходимо для дальнейших рассуждений в рамках эволюционной теории. Но чтобы построить хорошее древо, нужен опыт.
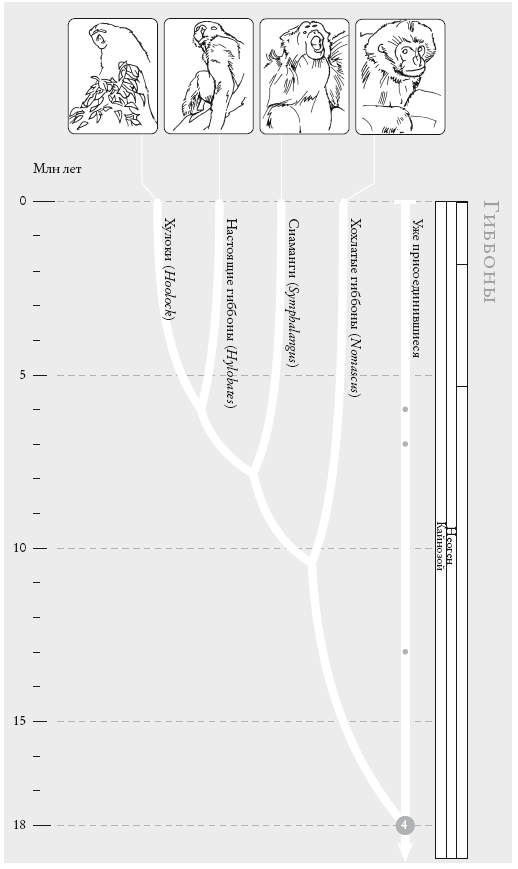
ГИББОНЫ. Двенадцать видов гиббонов сейчас объединяют в четыре группы. Порядок ветвления внутри них остается спорным (см. “Рассказ Гиббона”).
На рис. (слева направо): восточный хулок (Hoolock leuconedys), быстрый гиббон (Hylobatesagilis), сиаманг (Symphalangus syndactylus), желтощекий номаск (Nomascus gabriellae).




