



Алексей Ржешевский
Волосы. Иллюстрированное пособие для врачей, трихологов и парикмахеров
© Ржешевский А.В., Войтова М.С., Кабанов А.С., текст, 2024
© Самоукина А.А., Черникович В.Ю., иллюстрации, 2024
© Оформление. ООО «Издательство «Эксмо», 2024
Что такое волос и волосяной фолликул?
Наше тело практически полностью покрыто волосами, за исключением лишь некоторых частей, таких как ладони или подошвы ног. Но по сравнению с другими млекопитающими у нас меньше волос на туловище и конечностях.
Волосы человека бывают двух основных типов: более короткие и тонкие пушковые, встречающиеся на всем теле, и более длинные и толстые терминальные – это волосы на голове, ресницы, брови, лобковые волосы, волосы на груди и на животе. Доля волос того или иного типа зависит от возраста и пола человека. У детей волосы в основном пушковые. У взрослых людей – около 30 % (у женщин) и 90 % (у мужчин) – поверхности тела покрыто терминальными волосами; у взрослого человека они обычно бывают около 20-180 мкм в диаметре и вырастают до 90 см в длину[1].
Мы рождаемся примерно с пятью миллионами волосяных фолликулов, около ста тысяч из которых расположены на голове. Считается, что у взрослых людей (увы и ах!) не происходит образования новых фолликулов. Но те, которые у нас уже есть, очень пластичны, ведь один волосяной фолликул в течение своей жизни может производить несколько различных типов волосяных волокон:
• тонкие непигментированные волосы лануго – во время внутриутробного развития;
• короткие (в основном непигментированные) пушковые или тонкие пигментированные промежуточные волосы – в предпубертатном периоде;
• длинные толстые концевые стержни волос – у взрослых.
Что касается скорости роста волос, то она меняется в ходе старения человека. В среднем у нелысеющих людей они быстрее всего растут после 40 лет, а в возрасте от 50 до 70 лет толщина отдельных волокон достигает максимума[2].
Строение волоса
Каждый волос имеет стержень и корень. Стержень – это видимая часть, которая выступает из кожи. Корень волоса находится в ней самой, в волосяном фолликуле[3]. Он окружен защитным слоем – корневым влагалищем, – и покрыт внешним и внутренним слоями оболочки.
Стержень волоса состоит из трех слоев: кутикулы, коры и мозгового вещества[4]. Давайте посмотрим на его поперечное сечение и обсудим более подробно, что он из себя представляет.
Как было сказано выше, стержень волоса имеет три слоя (рис. 1.1 и 1.2).
• Внутреннее ядро, иначе говоря, мозговое вещество, которое в основном заполнено неупорядоченно расположенными нитевидными структурами.
• Мозговое вещество окружено корой, которая содержит большую часть массы волокон, состоящих в основном из белков кератина и структурных липидов.
Кора плотно упакована и содержит веретенообразные клетки. Они расположены параллельно оси волокна и имеют диаметр примерно 1–6 мкм и длину 50-100 мкм. Чтобы представить размер такой клетки, достаточно разделить миллиметр на 10 частей: одна из них как раз и будет равна длине клетки коры. Не такая уж она и маленькая!
Именно кора стержня определяет, какими будут волосы: прямыми или кудрявыми. Корковые клетки имеют три различные области: ортокортекс, паракортекс и мезокортекс. Если распределение в коре ортокортексов и паракортексов симметричное, то поперечное сечение волоса будет круглым, а макроскопически волос будет прямым. Несимметричное распределение кортикальных клеток создает овальную форму поперечного сечения и обеспечивает вьющуюся форму волоса (рис. 2).

Рисунок 1.1. Строение стержня волоса.
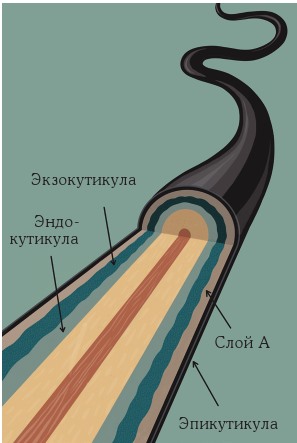
Рисунок 1.2. Строение стержня волоса.
• Снаружи находится единственный слой плоских клеток, составляющий кутикулу стержня – мертвые перекрывающиеся клетки. Они образуют защитный слой вокруг волоса. Эти клетки имеют толщину примерно 0,5 мкм и длину 45–60 мкм.
Кутикулярные клетки снаружи покрыты еще несколькими слоями (рис. 3):
• Самый внешний слой кутикулы называется эпикутикулой. Это липопротеиновая мембрана толщиной около 10–14 нм.
• Под ней находится слой А с высоким содержанием цистеина и толщиной 50-100 нм.
• Далее расположено экзокутикула, которая также содержит много цистеина, а ее толщина различается в диапазоне от 50 до 300 нм.
• Наконец, эндокутикула содержит мало цистеина и имеет толщину от 50 до 300 нм.

Рисунок 2. Сечение стержня волоса.

Рисунок 3. Тонкие структуры слоя кутикулы волоса.
Строение волосяного фолликула
Волосяной фолликул – это оболочка из кожи и соединительной ткани, окружающая корень волоса. Он начинается на поверхности эпидермиса – самого верхнего слоя кожи. Фолликулы, образующие пушковые волосы, заканчиваются у верхнего слоя дермы – следующего за эпидермисом слоя кожи. Фолликулы, образующие терминальные волосы, уходят намного глубже – внутрь дермы, а иногда даже в подкожный слой.
По структуре волосяной фолликул делится на верхнюю и нижнюю части. Обсудим особенности каждой из них.
Верхняя часть волосяного фолликула состоит из воронки и перешейка.
Воронка – это отверстие, из которого выходит волос. Она начинается на поверхности эпидермиса и продолжается до отверстия сального протока. Там находятся сальные железы, которые открываются в волосяные фолликулы. При стимуляции гормонами (такими как андрогены) они выделяют жирное кожное сало, которое защищает волосы и обеспечивает кожу водонепроницаемым барьером – своеобразной защитой. Отверстием сального протока воронка с помощью перешейка соединяется с выпуклостью – частью фолликула, к которой крепится мышца Arrector pili – выпрямитель волоса. На холоде эти мышцы сокращаются, и это заставляет волосы встать дыбом: по коже «идут мурашки». Выпуклость также содержит эпидермальные стволовые клетки. Собственно, выпуклость – это окончание постоянной области фолликула, которая не подвергается изменениям в зависимости от жизненного цикла волоса.
Нижний сегмент волосяного фолликула постоянно реконструируется при каждом новом цикле роста волос. Это настоящая фабрика стержня волоса. Нижний сегмент простирается от выпуклости до основания фолликула и состоит из луковицы и супрабульбарной области.
В волосяной луковице находятся волосяная матрица из быстро делящихся клеток – кератиноцитов, и дермальный сосочек с клетками соединительной ткани со специализированными клетками – фибробластами, а также кровеносные капилляры и нервные окончания. Дермальный сосочек снабжает кровью корень волоса. Он взаимодействует с матрицей, клетки которой имеют самую высокую скорость деления в организме, и именно поэтому они являются одними из самых уязвимых клеток для химиотерапии (вместе с клетками костного мозга, эпителия кишечника и собственно раковыми клетками).
Мигрирующие из выпуклости кератиноциты пролиферируют, образуя стержень растущего волоса. Чем больше таких делящихся кератиноцитов, тем больше и сама луковица, и диаметр стержня растущего волоса. Когда они перестают делиться, то превращаются в различные клетки волоса. Выпуклость и луковица разделены между собой длинным отрезком супрабульбарного (то есть «надлуковичного») эпителия[5][6][7][8].
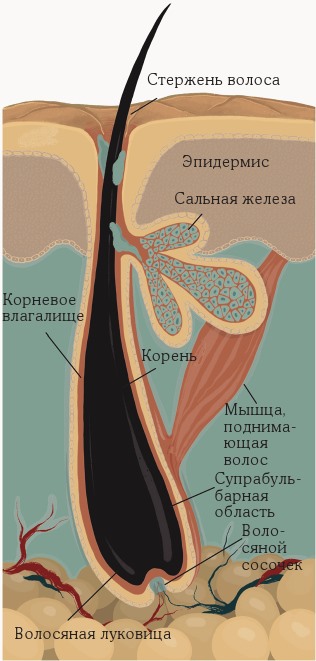
Рисунок 4. Сегменты волосяного фолликула.
Циклы жизни волосяного фолликула
Жизненный цикл волосяного фолликула делится на три основные фазы: анаген, катаген и телоген. В любой момент времени большинство фолликулов находятся в фазе анагена, остальные же пребывают либо в фазе катагена (2 % времени), либо в фазе телогена (10–15 % времени). Некоторые выделяют еще одну фазу – экзоген.
Поговорим о каждой из этих фаз подробнее.
Анаген
Волосы начинают свой жизненный путь в фазе анагена – это фаза активного деления клеток. В нижней части волоса клетки быстро делятся, и из волосяного фолликула растет новый стержень волоса. Эта фаза самая продолжительная, она длится от 2 до 6–7 лет. В любой момент времени около 90 % общего количества волос человека находится именно в анагене. В зависимости от того, в какой области тела растут волосы, фаза их роста будет длиться дольше или меньше, например, фаза роста волос на голове составляет несколько лет, поэтому они могут вырасти до метра в длину. Особенно короткой фаза анагена бывает у ресниц, бровей, волос в носу и ушах: они растут всего 100–150 дней, поэтому и не вырастают такими длинными.
В начале фазы анагена дермальный сосочек посылает сигнал эпителиальным стволовым клеткам в выпуклости. После стимуляции они мигрируют к основанию фолликула, и нижний его сегмент образует луковицу вокруг дермального сосочка. После этого он может сигнализировать клеткам матрикса о том, что они должны усиленно делиться, дифференцироваться и расти вверх, образуя новый волос.
Катаген
Фаза катагена – это короткий переходный период, инволюция или регресс. Она самая короткая из всех фаз и длится от двух до четырех недель. Деление клеток в матриксе прекращается, и нижний сегмент волосяного фолликула начинает регрессировать за счет запрограммированной гибели кератиноцитов матрикса. Стержень волоса отделяется от сосочка, а нижний волосяной фолликул превращается в эпителиальную нить, в результате чего дермальный сосочек перемещается вверх, чтобы снова войти в контакт с выпуклостью. Во время этого процесса образуется клубный волос с твердым белым узлом на конце.
Телоген
Фазу телогена можно назвать стадией покоя. Она длится около трех месяцев, и в это время не происходит значительного деления клеток или их гибели. Когда волосы полностью отделяются от сосочка, приток крови прекращается. Клубные мертвые волосы остаются на голове обычно около 100 дней. Они постепенно выталкиваются из кожи и со временем выпадают. После этого происходит процесс формирования нового волоса: у основания «пустого» волосяного фолликула начинают размножаться новые волосковые клетки, чтобы сформировать новый волос, и весь процесс начинается заново.
С каждым новым циклом продолжительность телогена увеличивается, поэтому длительность этого периода волосяных фолликулов влияет и на формирование волос: у стареющих животных этот процесс значительно замедляется. У людей синхронный жизненный цикл фолликула плода становится асинхронным вскоре после рождения, когда каждый волосяной фолликул начинает функционировать со своей собственной периодичностью.
В течение всей жизни фолликул реконструируется, поскольку рост волос не останавливается. Это особенно заметно при рассмотрении реакции волосяных фолликулов на андрогены: они вызывают увеличение волосяных фолликулов бороды у мальчиков-подростков и миниатюризацию волосяных фолликулов кожи головы у мужчин с андрогенной алопецией.
Волосы растут примерно на 1 см в месяц. Поскольку они постоянно переходят в фазу покоя, а затем выпадают, мы постоянно их теряем. У здорового взрослого человека за день может выпасть от 70 до 100 волос на голове, но при этом новые волосы постоянно растут и приходят на замену старым. Скорость выпадения может заметно увеличиться, если корни повредятся во время фазы роста или если много волос одновременно перейдут в фазу покоя. В том случае, если на какой-то области кожи новые волосы перестают расти и не заменяют старые, она станет лысой. Такой тип облысения называется алопецией. При некоторых типах алопеции волосы могут снова начать расти, но, к сожалению, облысение может быть и постоянным. Типичным примером является постепенное выпадение волос с возрастом у мужчин[9][10][11][12][13].
Волосы в эмбриональном периоде
Примерно на девятой неделе беременности эпидермис плода дает начало маленьким зачаткам специализированных клеток, которые образуют волосяной фолликул и связанные с ним придатки. Клетки будущей дермы подают сигнал эпидермальным клеткам о пролиферации и миграции вниз, к дерме. В ответ эпидермис дает указание нижележащим дермальным клеткам сбиваться в кучку и образовывать дермальный сосочек.
Первым признаком развития волосяного фолликула является формирование волосяной плакоды: по мере того как развивающийся фолликул расширяется вниз и обволакивает дермальный сосочек, клетки в его основании поддерживают высокую скорость деления. Во время созревания фолликула эти пролиферирующие (матричные) клетки начинают дифференцироваться во внутреннюю корневую оболочку (IRS), из которой потом развивается будущий стержень волоса. Внешний слой клеток становится внешней корневой оболочкой (ORS), которая прилегает к эпидермису и окружена снаружи базальной мембраной.
После этого активное деление клеток в матриксе прекращается, и нижние две трети волосяного фолликула быстро отмирают (в стадии катагена). Эпителиальная нить, окруженная базальной мембраной, тянет дермальный сосочек вверх, и он останавливается чуть ниже основания выпуклости (в стадии телогена). В первом цикле роста волос телоген длится примерно один день, однако в дальнейшем эта фаза становится все более длительной. Новый цикл регенерации волос (анаген) начинается с появления пролиферирующего зародыша волос, и его развитие с образованием зрелого фолликула имеет значительное сходство с развитием фолликулов у эмбриона.
После завершения процесса формирования фолликулов у плода имеется волосяной фолликул, содержащий матрикс (полученный из эктодермы) и нижележащий дермальный сосочек (полученный из мезодермы). Кроме того, вокруг каждого фолликула находятся арректорная мышца и сальная железа.
Когда формируется волосяной фолликул, у плода начинают расти волосы лануго. Они очень тонкие и короткие. В конечном итоге на сроке беременности примерно от 36 до 40 недель волосы лануго заменяются пушковыми волосами, покрывающими большую часть тела. Между тем на некоторых участках тела, например, на коже головы, бровях и ресницах, уже можно найти и более густые терминальные волоски.
После рождения человека новые волосяные фолликулы у него, увы, больше не образуются, а вот их размер меняться как раз может[14][15]. Циклы роста и дегенерации волос сменяют друг друга на протяжении всей жизни, что подтверждает существование стволовых клеток, поддерживающих эти процессы.
Гены циркадных ритмов и циклы роста волос
Наши суточные ритмы (время отхода ко сну, время пробуждения и максимальной работоспособности) регулируются не только внешними стимулами в виде смены дня и ночи. У всех видов живых организмов, имеющих суточные (циркадные) ритмы, включая животных, растения, грибы и даже цианобактерии, внутри имеется своеобразный биохимический «маятник»: белки, вступающие друг с другом в циклические реакции с периодичностью примерно одни сутки.
Биохимический «маятник» человека состоит из факторов транскрипции CLOCK и BMAL1 и генов-мишеней: трех генов с говорящим названием Periods (Per 1, 2 и 3) и двух криптохромов – Cryptochromes (Cry 1 и 2). Следует отметить, что криптохромы – это очень древние и эволюционно консервативные белки. Они присутствуют не только у животных, но и у растений, где выполняют функцию сенсоров света и тьмы и таким образом регулируют также циркадные ритмы[16].
CLOCK и BMAL1 активируют PER и CRY, а те в свою очередь ингибируют транскрипционную активность BMAL1-CLOCK, формируя петлю отрицательной обратной связи. Это приводит к ритмической экспрессии комплекса BMAL1-CLOCK с периодичностью 24 часа, и этот комплекс служит своеобразным биохимическим маятником.

Рисунок 5.1. Работа комплекса CLOCK/BMAL1 лежит в основе механизма циркадных ритмов млекопитающих. Он активирует несколько генов, среди которых Per 1, 2, 3 и Cry 1, 2. Белки – продукты этих генов – перемещаются в ядро, где ингибируют транскрипционную активность комплекса CLOCK/BMAL1. REV-ERBα – еще один ген, контролируемый комплексом CLOCK/BMAL1, белковым продуктом которого является отрицательный регулятор экспрессии Bmal1. Кроме того, REV-ERBα подавляет экспрессию ингибитора клеточного цикла – p21. В отсутствие BMAL1 подавление REV-ERBα приводит к высокой экспрессии p21 и задержке зародышевых клеток волос в фазе G1, что останавливает анаген.
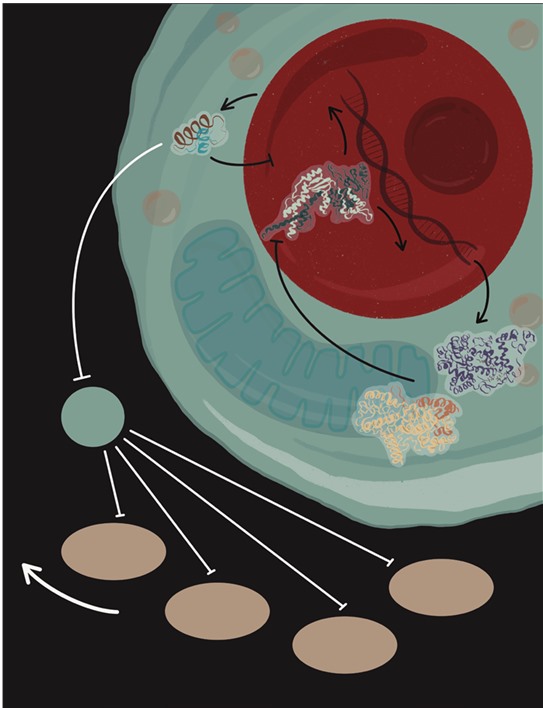
Рисунок 5.2. Обозначения: APM – мышца arrector pili, Bu – выпуклость, CH – клубный волос, CTS – соединительнотканная оболочка, DP – дермальный сосочек, EM – эпителиальная мембрана, HS – стержень волоса, IRS – внутреннее корневое влагалище, Ma – матрица, ORS – внешняя корневая оболочка, SB – сальная железа, SHG – вторичный росток волос[17].
Функции генов циркадных ритмов не исчерпываются только самими циркадными ритмами – они задействованы также и в росте волос. В телогене и раннем анагене активируются гены-мишени комплекса CLOCK/BMAL1. Из-за того что в коже они экспрессируются с четким циркадным ритмом, амплитуда уровня их экспрессии наиболее высока во время телогена и раннего анагена. Это указывает на то, что их активность зависит как от циркадного механизма, так и от цикла роста волос[18][19][20].
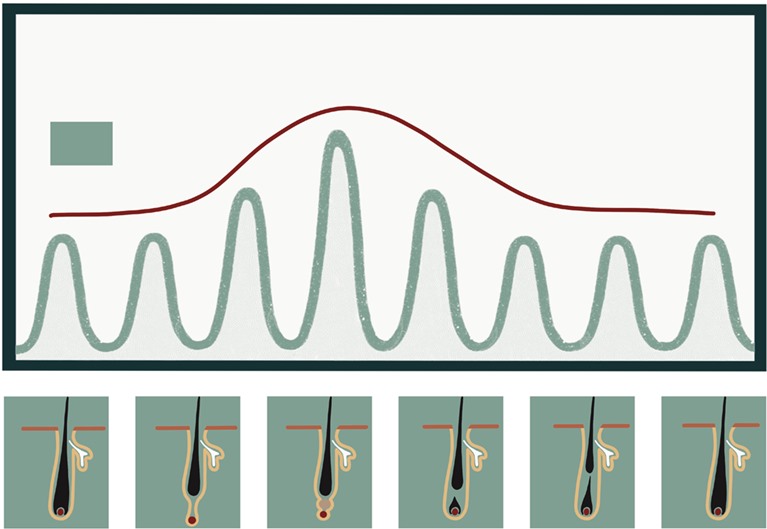
Рисунок 6. Диаграмма, отражающая ритмическую циркадную экспрессию гена Dbp (активатор транскрипции) на разных фазах цикла роста волос. Амплитуда экспрессии Dbp коррелирует с прогрессированием цикла волосяного фолликула: она становится максимальной во время телогена[21].
Гены циркадных часов экспрессируют все типы клеток кожи, но местом наиболее заметной ритмической экспрессии циркадных генов во время телогена и раннего анагена является вторичный зародыш волоса. Это отдел, расположенный между дермальным сосочком и выпуклостью. Он содержит пролиферативные стволовые клетки, которые мигрировали из выпуклости во время позднего катагена и раннего телогена[22][23][24]. Вторичные зародышевые клетки волоса во время инициации анагена активируются первичными, в результате чего формируются размножающиеся клетки волосяного матрикса, а в конечном итоге и стержень волоса[25][26][27].
У мышей, мутантных по генам Clock и Bmal1, анаген значительно задерживается, причем этот эффект наиболее выражен у мышей с дефицитом именно Bmal1. Такие мыши имеют целый ряд отличий от мышей дикого типа. Исследования показали следующее.
• В то время как в ходе анагена в большинстве волосяных фолликулов у здоровых мышей уже образовались волосяной матрикс и стержень волоса с волосяной луковицей, мутантные по Bmal1 животные все еще оставались в самой первой его фазе. В результате волосяные фолликулы экспериментальных животных возобновили нормальное развитие цикла роста волос лишь после почти недельной задержки. При этом аномалий в структуре зрелых фолликулов анагена у мутантных по Bmal1 или Clock мышей не было, что подтверждает участие генов циркадных часов, в первую очередь, в механизмах перехода телоген-анаген.
• Во вторичном зародыше волос раннего анагена в волосяных фолликулах мутантных по Bmal1 животных отсутствовали делящиеся клетки. При этом эпидермис и дерма этих мышей их содержали, значит, дефект пролиферации специфичен именно для волосяного фолликула[28][29].
• Во вторичном зародыше волос волосяных фолликулов мутантных животных отсутствовал фосфорилированный белок ретинобластомы (Rb) (маркер прохождения клеточного цикла через контрольную точку G1-S), хотя в норме его очень много. Это говорит о том, что в волосяных фолликулах экспериментальных мышей клетки-предшественники вторичного зародыша волос раннего анагена задерживаются в контрольной точке клеточного цикла G1-S.
• В коже мышей с дефицитом Bmal1 во время телогена оказалась затронута экспрессия нескольких генов-мишеней CLOCK-BMAL1, в том числе Rev-Erbα. Она снижалась в целых пятнадцать раз! Мы знаем, что REV-ERBα напрямую подавляет экспрессию гена, кодирующего ингибитор клеточного цикла G1-p21. Таким образом, в мутантной коже p21 активируется примерно в 2,5 раза сильнее.
• Активация p21 у экспериментальных животных привела к замедлению прогрессирования клеточного цикла в фазах G1-S в клетках-предшественниках вторичного зародыша волос[30].
Пигментация волос
Пигментация волос – это результат активности специальных клеток меланоцитов. Они названы так потому, что эти клетки производят пигмент меланин и откладывают его в стержень волоса по мере его формирования[31]. Меланоциты находятся в луковице волосяного фолликула.
Пигментация фолликулов чувствительна к многочисленным внутренним факторам. На нее влияют:
• фаза цикла роста волос;
• место расположения фолликула на теле;
• раса и пол;
• чувствительность к гормонам;
• генетические дефекты;
• возрастные изменения.
Пигментацию волос регулирует множество различных веществ, таких как факторы роста, цитокины, гормоны, нейропептиды и нейромедиаторы, эйкозаноиды, циклические нуклеотиды, питательные вещества, микроэлементы. Многие из этих веществ могут действовать через механизмы межклеточной сигнализации[32].