



Александр Козлов
Древнейшая история Пензенского края: мифы и реальность. Взгляд на историю с точки зрения новейших исследований ДНК-генеалогии
4. Мои предки осели в Поволжье в самом начале роста ветви Z1941, возможно, утратив связь с будущими финнами и карелами еще на рубеже нашей эры. Тем не менее, скорее всего, мой род берет начало именно в Карелии. И уже в исторические времена кто-то из него переселился в Среднее Поволжье. Со времен раннего Средневековья по Волге проходил оживленный торговый путь из Ирана и Средней Азии к Балтике и Северному Уралу, а потому столь дальние путешествия жителей бассейна Ладожского и Онежского озер вряд ли можно считать экзотикой. В этой связи интересен следующий факт: как только я узнал из персональной расшифровки результатов теста о возможных семейных «корнях» из Карелии, я запросил у близких родственников о такой возможности. Каково же было мое удивление, что в Карелии до сих пор живет моя родня по отцовской линии: ветвь родных старшего брата моего прадеда. Вот и не верь после этого в ДНК-генеалогию!
Общая же картина развития человечества представляется следующей. Анатомически современный человек неафриканского происхождения окончательно сформировался, видимо, в гигантском треугольнике, включающем территории от европейской Атлантики до Урала (возможно, и до южной Сибири) и, возможно, юг до северной Месопотамии (Леванта) – примерно 65–50 тыс. лет назад. Он оставил после себя верхнепалеолитические культуры, которые несколькими потоками распространились в Европу (с 45 тыс. лет назад), оттуда – предположительно в сторону Африки через Испанию; на Ближний Восток (с 40 тыс. лет назад) и в Прибайкалье (с 40 тыс. лет назад). Вторая волна расселения современного человека из Центральной Азии в Европу состоялась в интервале 20–9 тыс. лет назад (R1a) и 16–5 тыс. лет назад (R1b).
Эту картину дополняют самые последние данные по гаплогруппе костенковского человека с археологической датировкой 36–39 тыс. лет назад. Его гаплогруппа оказалась С-М130, и это самая древняя гаплогруппа из всех найденных ископаемых гаплогрупп в мире! Гаплогруппа С была обнаружена и в Испании (La Braña), но там был более недавний нисходящий субклад, да и датировка – «всего» 7960–7750 лет назад. Так что в упомянутом гигантском треугольнике происхождения человека, или, во всяком случае, европеоидов – Русская равнина занимает приоритетное место.
Можно считать твердо доказанными древние миграции предков нынешних восточных славян в Индию несколько тысячелетий назад. И современные этнические русские гаплогруппы R1a, и современные индусы той же гаплогруппы происходят от одних и тех же общих предков – ариев, которые жили примерно 5 тыс. лет назад.
Древние индийцы принципиально отличались от древних предков – современных европейцев. И это неудивительно – у тех в основном другой род, R1b. Он к ариям не имеет отношения. Действительно, при изучении 367 браминов из разных штатов Индии гаплогруппы R1b не было найдено ни у одного. Для сравнения: у браминов Западной Бенгалии доля гаплогруппы R1a – 72 %, у браминов штата Уттар Прадеш – 68 %.
А сейчас 60 % мужского населения Европы – мы уже это отмечали – это гаплогруппа R1b. По современному языку – «индоевропейцы». Но это сейчас. А мы говорим про времена 6000–4000 лет назад. Тогда действительно «индоевропейцами» были только арии, они этот индоевропейский, а на самом деле арийский язык принесли и в Индию, и в Иран, и в Митанни. Не современные «индоевропейцы» принесли и не их предки в гаплогруппе R1b, они тогда в Индии не были. А принесли древние «индоевропейцы», носители гаплогруппы R1a, тогдашние арии.
Дело в том, что носители гаплогруппы R1a появились на Русской равнине примерно 4800 лет назад (прямой расчет дает 4600 лет назад, но есть некоторые ветви, добавляющие 200–300 лет), и они разделились на три (по меньшей мере) миграционных потока восточного и южного направления – один на Южный Урал (прибыл туда примерно 4000 лет назад, в самом начале II тыс. до н. э.), другой – на Кавказ и в Закавказье (Анатолия, Митанни), в интервале 4200–3600 лет назад, третий – в Среднюю Азию, и стал авестийскими ариями, тоже примерно 4000 лет назад. Те, кто прибыл на Южный Урал, тоже разошлись: одни ушли на юг, в Индию – и стали легендарными ариями, другие пошли дальше на восток и стали скифами. По сути, это одни и те же люди.
Арии – это носители гаплогруппы R1a в III–II тыс. до н. э. (c расширением на несколько тысячелетий в древность и на тысячелетие «вверх», до начала нашей эры, хотя эти границы условны), которые продвинулись с Русской равнины в Индостан, на Иранское плато, и через Кавказ в Анатолию, Митанни и на Аравийский полуостров, пронеся по всем этим направлениям и регионам свою гаплогруппу R1a. Более того, это они же, арии, прошли далеко за Урал, на тысячи километров восточнее, и ту же гаплогруппу R1a и те же гаплотипы определяют в ископаемых костях в Минусинской котловине, между Кузнецким Алатау на севере и Восточными и Западными Саянами на юге, в местах так называемого скифского круга.
Именно эта гаплогруппа R1a, в виде совершенно определенной метки в Y‐хромосоме ДНК и передающаяся по наследству вплоть до настоящего времени, является идентификатором ариев и их потомков у современных славян – и в Индии, и в Иране, и в Месопотамии, и на Ближнем Востоке, и на Алтае, у скифов, и вообще везде, куда они ни прибывали.
А вот как Анатолий Клёсов описывает путь развития второй по численности у русских и родной для меня гаплогруппы N: «Поставим гаплогруппу NO (предковая для N) и затем N и ее основные субклады в хронологические рамки, как мы делаем здесь в отношении всех гаплогрупп. Гаплогруппа NO образовалась 287 снип-мутаций назад, то есть 41 300 лет назад, путем соответствующей мутации (превращение тимина в цитозин в нуклеотиде Y-хромосомы под № 13 360 045) и далее, 250 мутаций назад, или 36 тыс. лет назад, образовалась гаплогруппа N. Произошло это, вероятно, в Юго-Восточной Азии, как вариант рассматривается Вьетнам, и оттуда носители гаплогруппы N постепенно смещались по территории будущего Китая на север, к Алтаю. Это видно по шлейфу мутаций в Y-хромосомах современных носителей гаплогруппы N. Ископаемых ДНК на этом миграционном потоке пока нет, но, когда они появятся, это позволит уточнить, каким путем люди прошли из Юго-Восточной Азии на северо-запад Европы, какие археологические культуры оставили на своем пути, на каких языках они говорили».
На территории Китая сейчас выявлены снип-мутации (у современных китайцев), которые образовались 152 снипа, или 22 тыс. лет, назад; 120 снипов, или 17 300 лет, назад (субклад N1a-L729); 109 снипов, или 15 700 лет, назад. Это может свидетельствовать о древних корнях гаплогруппы N в Китае. Это субклады Z4762, L729, Z1956, переходные между N и N1a. Далее гаплогруппа N появляется на Алтае, и ее субклад (F1419), образованный 76 снип-мутаций, или 10 900 лет, назад, обнаружен у современного жителя Хакасии. Этот субклад – первый нижестоящий от N1a1-M46, который образовался 102 снип-мутации, или 14 700 лет, назад. Следующим идет субклад L708, который образовался 72 снипа, или 10 400 лет, назад, и его уже находят у башкир, коми, мордвы, татар. Субклад N1a1 есть почти у всех якутов, и далее он сопровождает все дальнейшие миграции носителей гаплогруппы N через Урал и до Балтики, до скандинавских стран – Финляндии, Норвегии, Дании, Швеции, а также вдоль южного берега Балтики – у эстонцев, латышей, литовцев, поляков, немцев. Гаплогруппа Na1a обильно представлена по всему уральскому миграционному пути и маркирует выход с Урала на Русскую равнину и далее на запад. Больше всего ее у удмуртов – 67 %, то есть две трети от всего мужского населения. У марийцев – 61 %, у чувашей – 28 %, у башкир – 17 %, у татар – 16 %, у мордвов – 15 %. То есть мы уже выходим на уровень представительства гаплогруппы N1a1 у этнических русских – в среднем 14 %. Поскольку носители гаплогруппы N прошли на север, то у коми, например, ее уже 51 %.
Гаплогруппа N1a1-M46 (с датировкой образования 102 снипа, или 14 700 лет, назад) определенно образовалась еще в Южной Сибири, как и ее нижестоящие субклады, отстоящие более чем на 50 снипов (7200 лет назад) от настоящего времени. Это субклады F1419 (76 снипов назад) и L708 (72 снипа назад). Передвижения на запад через Урал заняли тысячелетия и оставили характерный след в виде пермских языков, распространенных в России на территории Удмуртии, Республики Коми и Пермского края. Это уральские языки, которые входят в состав финно-угорской ветви. Мы уже видели выше, что в Удмуртии две трети мужского населения имеют гаплогруппу N. Видимо, там, на Урале, образовался субклад N1a1-L1026 (43 снипа, или 6200 лет, назад) и далее VL29 (29 снипов, или 4200 лет, назад). Это уже времена – или близкие к ним – выхода носителей гаплогруппы N1a1 с Урала на Русскую равнину.
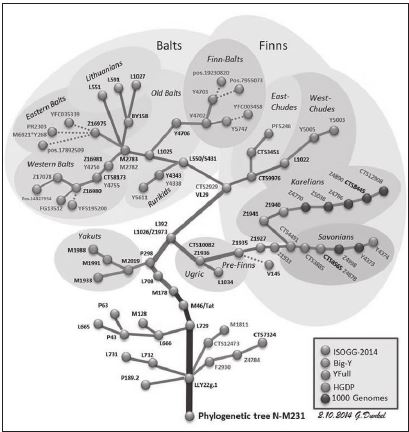
Весьма наглядно путь развития гаплогруппы N представлен на ее Филогенетическом дереве (находится в свободном доступе на Yandex.ru)
Судя по серьезным различиям в картине мутаций гаплотипов финнов и южных балтов, эти народности, или племена, разошлись еще на Урале не менее чем за 4000 лет до их прибытия на Балтику. Похоже, что разошлись они от субклада N1a1-L1026, который образовался 43 снипа, или 6200 лет, назад. Финны сохранили свой финно-угорский язык и прибыли на территории будущей Финляндии не ранее начала нашей эры, возможно, и к середине I тыс. н. э. Будущие бал-ты – литовцы и латыши прибыли на свои будущие земли несколько ранее, чем финны, – в конце прошлой эры или в самом начале новой эры. За время перехода с Урала они утратили свой угорский язык и перешли на индоевропейский. Видимо, это произошло под влиянием носителей гаплогруппы R1a, потомков фатьяновской культуры (4300–3500 лет назад), по территориям которой шла миграция будущих балтов. В настоящее время 40 % литовских и латышских мужчин имеют гаплогруппу N1a1 и столько же – гаплогруппу R1a, остальные гаплогруппы занимают единицы или доли процентов, кроме 12 % гаплогруппы R1b у латышей (у литовцев ее 5 %). Эстония занимает место в составе будущих южных балтов и примерно 2000–1500 лет назад в составе будущих финнов. Дальнейшее продвижение их на запад происходило еще позже, и на территории будущих Швеции, Норвегии, Дании гаплогруппа N пришла уже совсем недавно (по историческим меркам). Все гаплотипы там либо «финские», либо «южнобалтийские», фактически балтославянские. Более того, там их мало. В современной Швеции носителей гаплогруппы N – 7 %, в Норвегии – 2,5 %, в Дании – 1 %. Поэтому все рассуждения поклонников «норманской теории», что гаплотипы русских князей, членов Российского дворянского собрания, «похожи на шведские», могут вызывать только недоумение и смех. Очень уж хотят «норманисты» видеть шведов и прочих скандинавов в основе создателей Древней Руси, ее государственности! А на самом деле в Швеции две трети гаплотипов группы N1a1 имеют такой же вид, как и южнобалтийские, по сути, славянские, а остальная треть – как финские. «Шведского» там вообще ничего нет!
Приведу еще и общее дерево гаплогрупп современного человечества. На нем можно найти гаплогруппу N и гаплогруппу R1a и убедиться, что они на самом деле отстоят друг от друга довольно далеко.
А теперь еще раз перейдем к гаплогруппе N, рассмотрев ее в несколько больших деталях (см. рис).
Ну и в довершение этой главы нелишне будет показать Диаграмму эволюции гаплогрупп современного человечества и таблицу всех имеющихся гаплогрупп с датировками их образования. Они наглядно иллюстрируют то, как и когда образовывались и видоизменялись гаплогруппы на планете Земля.

Ствол гаплогрупп человечества

Гаплогруппа N в деталях

Диаграмма зволюции гаплогрупп современного человечестеа. На горизонтальной оси – основные гаплогруты Y-хромосомы чеюеечестеа, на вертикальной – абсолютная шкала времени. Общий предок альфа-гаплогруппы жил примерно 160 тыс. лет назад, общий предок бета-гаплогруппы (или гаплогрупп отВ до Т) – 64±6 тыс. лет назад (из статьи А.А. Klyosov and I.L. Rozhanskii, Advances in Anthropology, 2012). На диаграмме не показаны архаичные африканские линии АОО и АО (последняя сейчас заменила в номенклатуре линии Alb на диаграмме слееа), обновленное дерево гаплогрупп будет показано ниже
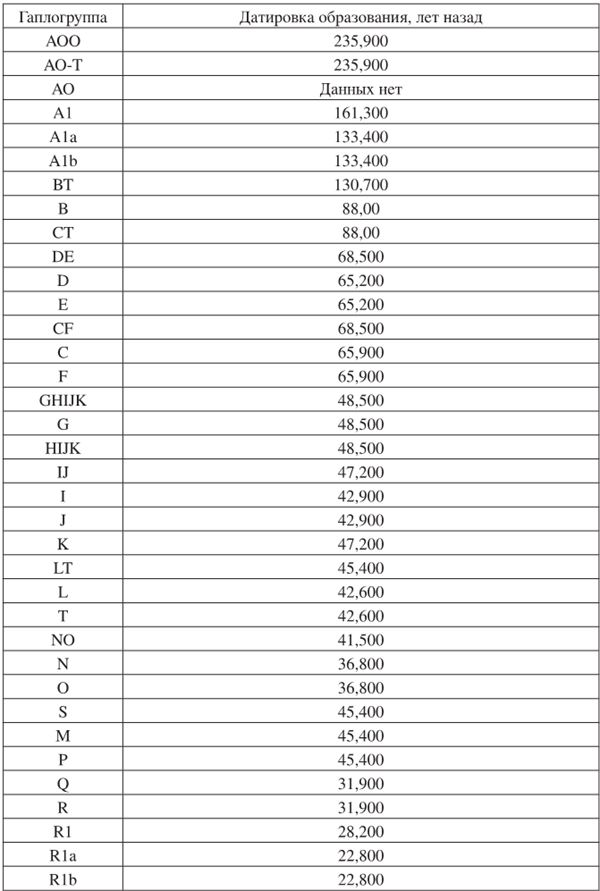
Таблица гаплогрупп с датировками образования
Эта таблица по данным расчетов группы Yfull (кто захочет, легко найдет о ней информацию в Интернете по адресу https://www.yfull.com/tree).
При рассмотрении датировок следует учесть, что для гаплогрупп ВТ, В и СТ, то есть первых в ряду «неафриканских», датировки показывают времена образования гаплогрупп и не учитывают прохождение гаплогруппой «бутылочного горлышка» популяции, которое произошло (или завершилось) 64 ± 6 тыс. лет назад. Все последующие гаплогруппы образовывались уже после прохождения гаплогруппой ВТ «бутылочного горлышка».
Здесь, очевидно, нелишне будет напомнить читателю, что это за зверь такой – «бутылочное горлышко» эволюции? Поэтому расшифровываю: эффект бутылочного горлышка – это сокращение генофонда (то есть генетического разнообразия) популяции вследствие прохождения периода, во время которого по различным причинам происходит критическое уменьшение её численности, в дальнейшем восстановленное. Сокращение генетического разнообразия приводит к изменению относительных и абсолютных частот аллелей генов, поэтому данный эффект рассматривается в числе факторов эволюции. Название «эффект бутылочного горлышка» наглядно демонстрирует один из способов отображения численности популяции. Если схематически изобразить численность популяции в один момент времени в виде горизонтальной полоски или эллипса, а численность в последующие моменты – таким же образом, но на пропорциональную величину выше над первым изображением, то случаи резкого снижения численности будут выглядеть как сужения рисунка в верхней части, что графически подобно горлышку бутылки.
Еще немного о ДНК-генеалогии
Для закрепления материала и более глубокого освоения вновь открывшейся нам науки ДНК-генеалогии вспомним ее основные положения и определения.
ДНК-генеалогия – одна из областей естествознания, молекулярная история. Она рассматривает закономерности наследования изменений нерекомбинантных (негенных) участков ДНК человека в ходе его эволюции на шкале времени от десятков и сотен лет до миллионов лет. Другими словами, ДНК-генеалогия изучает динамику накопления мутаций в ДНК человека, используя подходы химической и биологической кинетики, которые в свою очередь являются частью физической химии. Важнейшая особенность методологии ДНК-генеалогии – определение констант скоростей мутаций в ДНК (в первую очередь в тандемных повторяющихся последовательностях Y-хромосомы, так называемых маркерах, которых, по оценкам, имеется примерно 2500, а также накопления снипов (необратимых мутаций в ДНК) и приложение этих констант к расчетам хронологии древних событий – древних миграций человека, времен жизни общих предков изучаемых популяций.
Впервые термин ДНК-генеалогия был использован в 1992 году в работе японских авторов, но в виде «митохондриальная ДНК-генеалогия». С тех пор авторы этот термин не использовали. Когда Анатолий Клёсов в 2006 году ввел этот термин в сочетании «ДНК-генеалогия» для изучения Y-хромосомы, попгенетики подняли в сети страшный шум, уверяя, что это плохое название, что его никто не использует и что оно никогда не приживется. Но прижилось! И теперь ввод словосочетания «ДНК-генеалогия» в поисковую систему Яндекс дает 7 тыс. результатов, а в Google – на русском языке – 252 тысячи результатов. Еще больше эти цифры при вводе словосочетания «Академия ДНК-генеалогия»: соответственно, 4 млн результатов – на Яндекс, 126 тыс. результатов – в Google. Причем с каждым годом это количество растет.
Вот как сам Анатолий Клёсов определяет суть основополагающих понятий ДНК-генеалогии, и в частности ее прикладной науки – молекулярной истории: «Суть понятия молекулярной истории в том, что становится возможным следить за передвижениями древних народов не с помощью лопаты и кисточки археолога, не обмеряя черепа, не хитроумно расплетая созвучия и значения слов в живых и мертвых языках, не изучая древние фолианты в библиотеках и монастырях, а прослеживая за метками в Y-хромосомах наших ДНК. Они, эти метки, не могут «ассимилироваться», или «поглотиться», другими языками, культурами или народами, как это происходит тысячелетиями с языками, культурами, народами в рамках понятий истории, лингвистики, этнографии, антропологии. Иначе говоря, методология новой исторической науки, «молекулярной истории», или ДНК-генеалогии, основывается на изучении молекул нуклеиновых кислот, а именно ДНК, дезоксирибонуклеиновой кислоты, в человеческих организмах, как живых, так и в древних костных останках. То, что еще несколько лет назад казалось каким-то развлечением, оказалось, дает истории, антропологии, археологии, лингвистике возможность проверить концепцию, рассмотреть данные под принципиально другим углом, связать воедино, казалось бы, разрозненные части общей картины наших знаний об окружающем мире».
Разумеется, не следует понимать буквально эти слова Анатолия Алексеевича Клесова так, что отныне не нужно читать фолианты, изучать языки и проводить археологических раскопок. Напротив, это все приобретает еще большую значимость и ценность, поскольку усиливает выводы традиционных подходов в совокупности с выводами ДНК-генеалогии при условии взаимного их согласования. Или, напротив, заставляет пересматривать, казалось бы, устоявшиеся выводы, если они входят в явное противоречие с данными ДНК-генеалогии. Поэтому лично я очень внимательно слежу за всеми современными открытиями наших (и не наших в том числе) археологов. А в последнее время еще и за открытиями специалистов в области тестирования и изучения древних ДНК. Но читатель вправе сам определять свои приоритеты в методике изучения истории. Не нравится ДНК-генеалогия – не изучай. Воспользуйся традиционными методами и практиками. Но знай, что теперь уже практически любые серьезные исторические исследования не обходятся без анализа имеющихся мировых баз исследованных гаплогрупп, как современных жителей человечества, так и всевозможных останков, выявленных в ходе археологических раскопок (так называемые – палео-ДНК).
При этом только не надо путать ДНК-генеалогию с попгенетикой! Чем популяционная генетика отличается от ДНК-генеалогии? Да практически всем. Адресуемыми вопросами, целями и задачами, методологией, расчетным аппаратом, характером выводов, определениями популяций. Но, главное, это вообще другая область науки. Популяционная генетика – описательная, по сути, а ДНК-генеалогия – аналитическая. Попгенетика описывает структуру популяций, ДНК-генеалогия описывает историю популяций. Название о многом говорит. Не случайно биологическая химия – это химия, а химическая физика – это физика. Физическая химия – это химия. Биофизика, то есть биологическая физика – это физика. Второе слово в таких составных названиях дисциплин показывает, к какой области науки дисциплина относится. Популяционная генетика – это генетика. А ДНК-генеалогия к генетике, по сути дела, никакого отношения не имеет, это – генеалогия, то есть история.
Так вот еще раз о популяционной генетике. Популяционная генетика – это часть генетики, о чем говорит ее название, но со своими особенностями. Главная задача популяционной генетики – это выявление связи между генотипом и фенотипом, то, к чему ДНК-генеалогия не имеет отношения. Популяционная генетика иногда тоже рассматривает гаплогруппы и гаплотипы Y-хромосомы, как и ДНК-генеалогия, но на этом сходство заканчивается. Аппарат популяционной генетики, применяемый для интерпретации получаемых данных, например, «метод главных компонент», характеризуется со стороны ДНК-генеалогии как совершенно примитивный и ведущий к заблуждениям, и не используется в ДНК-генеалогии. Он не отвечает задачам ДНК-генеалогии.
В целом же генетика человека – это наука, изучающая наследственность и изменчивость признаков, определяющих врожденные особенности человека, и передаваемых, как правило, через гены. Но ДНК только на 2 % состоит из генов, а в Y-хромосоме генов вообще ничтожная доля. ДНК-генеалогия в настоящее время изучает в основном Y-хромосому, и не изучает наследственность и изменчивость упомянутых признаков, передаваемых через гены. Но дело даже не столько в этом. Направления и области науки определяются не объектами исследования, а методологией исследования. Именно методология исследования отличает, например, химию от физики, хотя объекты часто одинаковы. Методология генетики совершенно другая, чем методология ДНК-генеалогии. У них разные задачи исследований. ДНК-генеалогия, по сути, историческая наука, она оперирует хронологией, датировками древних событий, и для этого ДНК-генеалогия использует свой расчетный аппарат, которого нет в методологии генетики. ДНК-генеалогия использует и применяет методы физико-химической кинетики в отношении к ДНК, переводит картину мутаций в хронологические, исторические показатели, как указывалось выше. Это вообще не входит в методологию генетики, у нее другой экспериментальный и аппаратурный базис.
Итак, повторяем и закрепляем изученные истины: экспериментальные данные ДНК-генеалогии – это картина мутаций в нерекомбинантных (то есть не комбинирующихся с другими в ходе передачи наследственной информации потомкам) участках мужской половой хромосомы (на самом деле – и в митохондриальной ДНК, но пока в малой степени, так как информативность мтДНК в ДНК-генеалогии в настоящее время несравненно меньше, чем информативность Y-хромосомы). Речь здесь идет об информативности для исторических исследований, которую определяет картина мутаций как в Y-хромосомах отдельных людей, так и их групп, племен, родов, популяций. Естественно, в данном контексте термин «популяций» относится к мужской половине рассматриваемой популяции. В ДНК-генеалогии популяция – это мужская часть населения, на которую распространяются выводы, полученные при рассмотрении определенной выборки. Популяция может или ограничиваться выборкой, или чаще быть шире выборки, но при этом подразумевается или доказывается, что выявленные закономерности выборки распространяются на рассматриваемую популяцию. Например, если в пределах популяции сделаны несколько независимых выборок, и результаты в пределах погрешности расчетов совпали, то они могут распространяться на всю популяцию, пока не найдено обратного.
Методология новой науки ДНК-генеалогии – перевод динамической картины мутаций в хронологические показатели, во времена жизни общих предков популяций, а на самом деле – общих предков древних родов и племен. То есть фактически по мутациям в ДНК производится расчет времен, когда в древности жили эти роды и племена, и как эти времена меняются от территории к территории, что может указывать на направления древних миграций. Используя эти замечательные возможности новой науки, мы и проследили миграционные потоки нужных и близких нам гаплогрупп (R1a и N1), и связали их с историей родного нам края и земель.
А теперь мы более подробно поговорим непосредственно о нашем родном крае, только теперь уже в нашем веке. И поможет нам в этом замечательный ученый, краевед, подвижник Павел Корнилович Шиндин. Его книга «Отчий край Николаевский район» – настоявший шедевр, подарок для жителей нашего города и области. История соседнего Николаевского, но в том числе и Кузнецкого края предстает в ней, что называется, в «полной красе». Мне остается только коротко и популярно изложить самое интересное.




